

Avian cone photoreceptors tile the retina as five independent, self-organizing mosaics
- PMID: 20126550
- PMCID: PMC2813877
- DOI: 10.1371/journal.pone.0008992
Avian cone photoreceptors tile the retina as five independent, self-organizing mosaics
Abstract
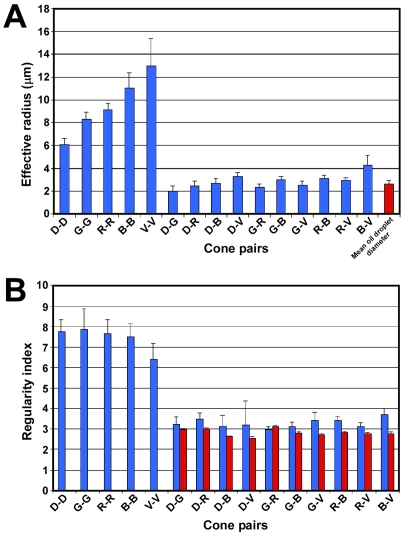
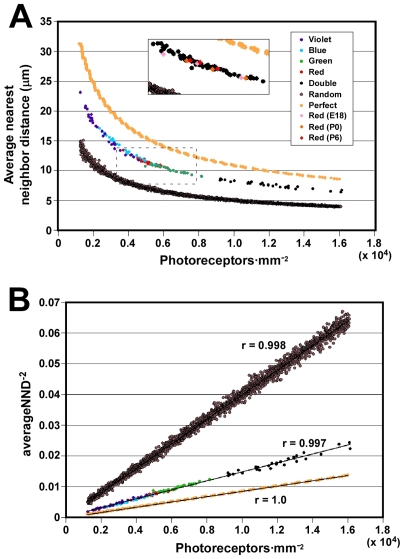
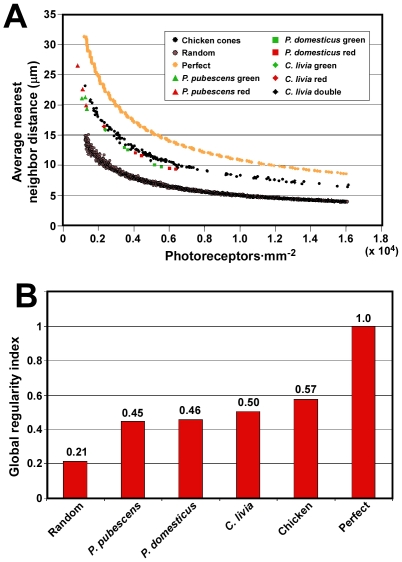
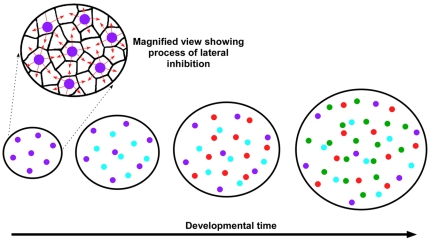
The avian retina possesses one of the most sophisticated cone photoreceptor systems among vertebrates. Birds have five types of cones including four single cones, which support tetrachromatic color vision and a double cone, which is thought to mediate achromatic motion perception. Despite this richness, very little is known about the spatial organization of avian cones and its adaptive significance. Here we show that the five cone types of the chicken independently tile the retina as highly ordered mosaics with a characteristic spacing between cones of the same type. Measures of topological order indicate that double cones are more highly ordered than single cones, possibly reflecting their posited role in motion detection. Although cones show spacing interactions that are cell type-specific, all cone types use the same density-dependent yardstick to measure intercone distance. We propose a simple developmental model that can account for these observations. We also show that a single parameter, the global regularity index, defines the regularity of all five cone mosaics. Lastly, we demonstrate similar cone distributions in three additional avian species, suggesting that these patterning principles are universal among birds. Since regular photoreceptor spacing is critical for uniform sampling of visual space, the cone mosaics of the avian retina represent an elegant example of the emergence of adaptive global patterning secondary to simple local interactions between individual photoreceptors. Our results indicate that the evolutionary pressures that gave rise to the avian retina's various adaptations for enhanced color discrimination also acted to fine-tune its spatial sampling of color and luminance.
Conflict of interest statement
Figures
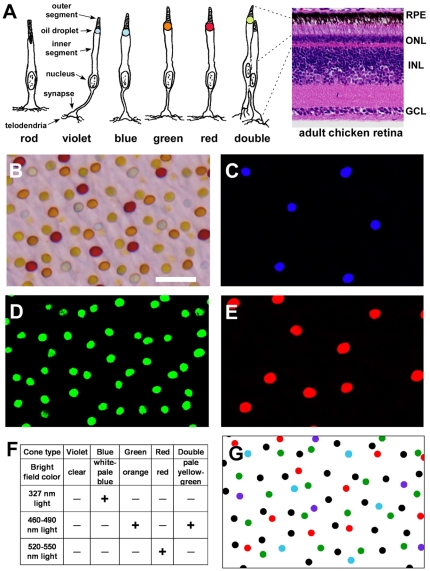
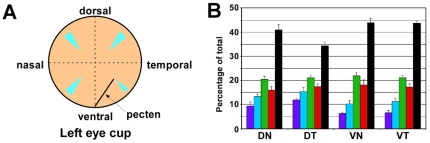
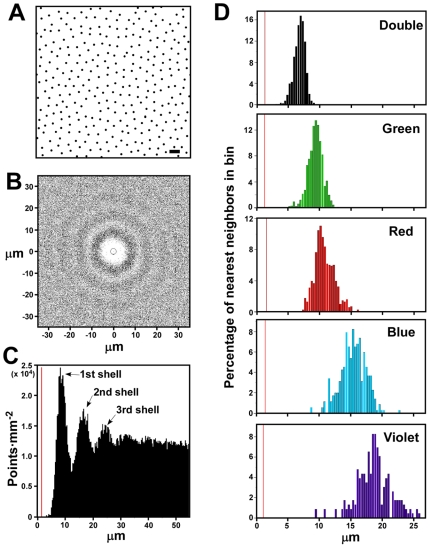
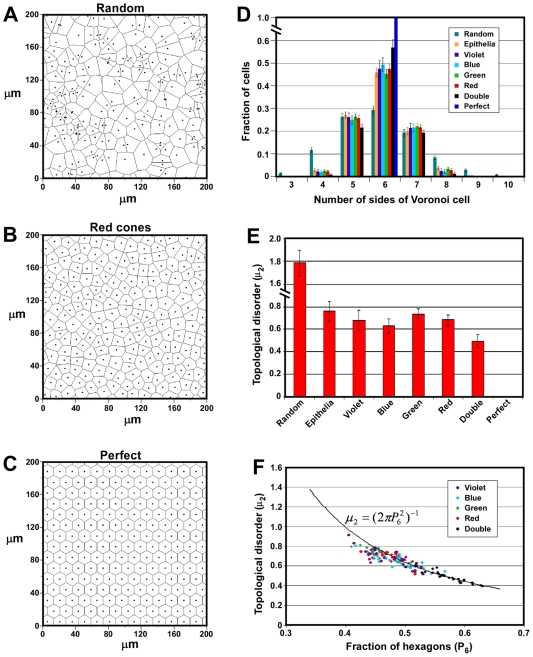
References
-
- Hart NS. Variations in cone photoreceptor abundance and the visual ecology of birds. J Comp Physiol A. 2001;187:685–697. - PubMed
-
- Bowmaker JK, Knowles A. The visual pigments and oil droplets of the chicken retina. Vision Res. 1977;17:755–764. - PubMed
-
- Maier EJ, Bowmaker JK. Colour vision in the passeriform bird, Leothrix lutea: correlation of visual pigment absorbance and oil droplet transmission with spectral sensitivity. J Comp Physiol A. 1993;172:295–301.
-
- Campenhausen M, Kirschfeld K. Spectral sensitivity of the accessory optic system of the pigeon. J Comp Physiol A. 1998;183:1–6.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
