

Chlorophyll-deficient mutants of Chlamydomonas reinhardtii that accumulate magnesium protoporphyrin IX
- PMID: 20127142
- PMCID: PMC2837180
- DOI: 10.1007/s11103-010-9604-9
Chlorophyll-deficient mutants of Chlamydomonas reinhardtii that accumulate magnesium protoporphyrin IX
Abstract
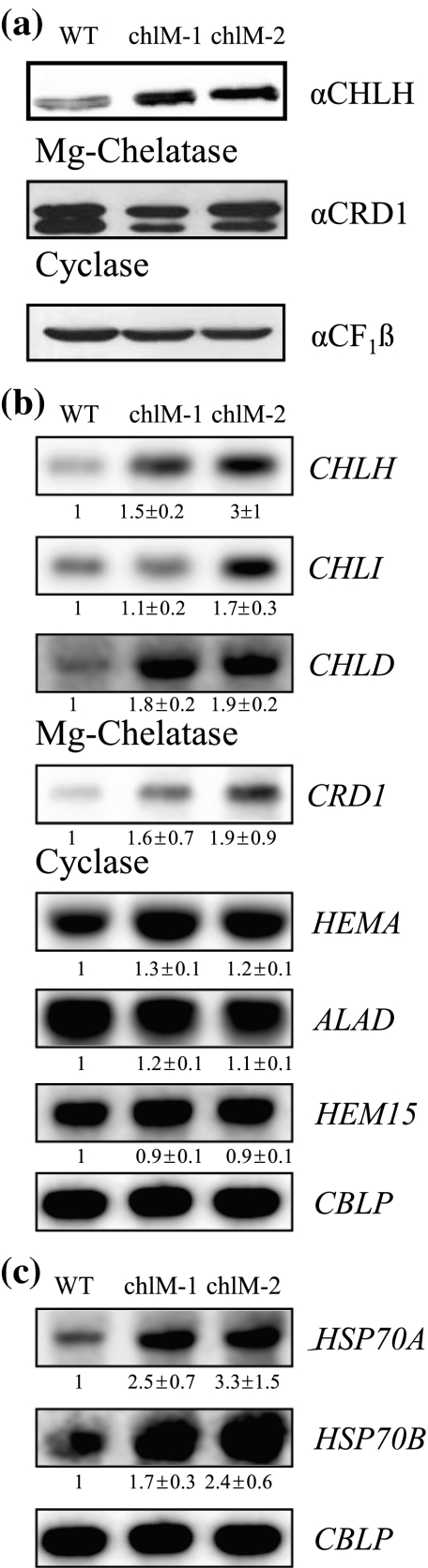
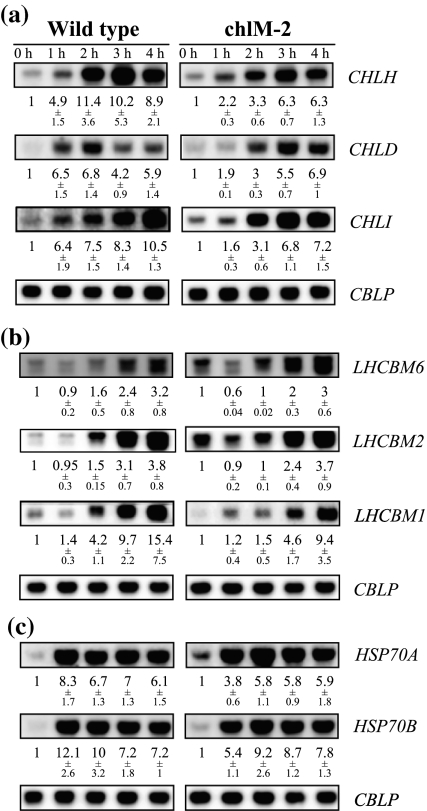
Two Chlamydomonas reinhardtii mutants defective in CHLM encoding Mg-protoporphyrin IX methyltransferase (MgPMT) were identified. The mutants, one with a missense mutation (chlM-1) and a second mutant with a splicing defect (chlM-2), do not accumulate chlorophyll, are yellow in the dark and dim light, and their growth is inhibited at higher light intensities. They accumulate Mg-protoporphyrin IX (MgProto), the substrate of MgPMT and this may be the cause for their light sensitivity. In the dark, both mutants showed a drastic reduction in the amounts of core proteins of photosystems I and II and light-harvesting chlorophyll a/b-binding proteins. However, LHC mRNAs accumulated above wild-type levels. The accumulation of the transcripts of the LHC and other genes that were expressed at higher levels in the mutants during dark incubation was attenuated in the initial phase of light exposure. No regulatory effects of the constitutively 7- to 18-fold increased MgProto levels on gene expression were detected, supporting previous results in which MgProto and heme in Chlamydomonas were assigned roles as second messengers only in the transient activation of genes by light.
Figures






Similar articles
-
Mg-protoporphyrin IX and heme control HEMA, the gene encoding the first specific step of tetrapyrrole biosynthesis, in Chlamydomonas reinhardtii.Eukaryot Cell. 2005 Oct;4(10):1620-8. doi: 10.1128/EC.4.10.1620-1628.2005. Eukaryot Cell. 2005. PMID: 16215169 Free PMC article.
-
Heme, a plastid-derived regulator of nuclear gene expression in Chlamydomonas.Plant Cell. 2008 Mar;20(3):552-67. doi: 10.1105/tpc.107.054650. Epub 2008 Mar 25. Plant Cell. 2008. PMID: 18364467 Free PMC article.
-
Phototropin involvement in the expression of genes encoding chlorophyll and carotenoid biosynthesis enzymes and LHC apoproteins in Chlamydomonas reinhardtii.Plant J. 2006 Oct;48(1):1-16. doi: 10.1111/j.1365-313X.2006.02852.x. Plant J. 2006. PMID: 16972865
-
Identification of Lhcb gene family encoding the light-harvesting chlorophyll-a/b proteins of photosystem II in Chlamydomonas reinhardtii.Plant Cell Physiol. 2001 Aug;42(8):849-56. doi: 10.1093/pcp/pce115. Plant Cell Physiol. 2001. PMID: 11522911
-
Functional Accumulation of Antenna Proteins in Chlorophyll b-Less Mutants of Chlamydomonas reinhardtii.Mol Plant. 2017 Jan 9;10(1):115-130. doi: 10.1016/j.molp.2016.10.001. Epub 2016 Oct 11. Mol Plant. 2017. PMID: 27742488
Cited by
-
Tetrapyrrole Metabolism in Arabidopsis thaliana.Arabidopsis Book. 2011;9:e0145. doi: 10.1199/tab.0145. Epub 2011 Jul 31. Arabidopsis Book. 2011. PMID: 22303270 Free PMC article.
-
Physiological, Cytological, and Transcriptomic Analysis of Magnesium Protoporphyrin IX Methyltransferase Mutant Reveal Complex Genetic Regulatory Network Linking Chlorophyll Synthesis and Chloroplast Development in Rice.Plants (Basel). 2023 Nov 6;12(21):3785. doi: 10.3390/plants12213785. Plants (Basel). 2023. PMID: 37960141 Free PMC article.
-
Mutagenesis selection and large-scale cultivation of non-green Chlamydomonas reinhardtii for food applications.Front Nutr. 2024 Sep 25;11:1456230. doi: 10.3389/fnut.2024.1456230. eCollection 2024. Front Nutr. 2024. PMID: 39385786 Free PMC article.
-
Tetrapyrrole Signaling in Plants.Front Plant Sci. 2016 Oct 19;7:1586. doi: 10.3389/fpls.2016.01586. eCollection 2016. Front Plant Sci. 2016. PMID: 27807442 Free PMC article. Review.
-
Systematic identification and characterization of genes in the regulation and biogenesis of photosynthetic machinery.Cell. 2023 Dec 7;186(25):5638-5655.e25. doi: 10.1016/j.cell.2023.11.007. Cell. 2023. PMID: 38065083 Free PMC article.
References
-
- Beale SI. Enzymes of chlorophyll biosynthesis. Photosynth Res. 1999;60:43–73. doi: 10.1023/A:1006297731456. - DOI
-
- Beck CF, Grimm B. Involvement of tetrapyrroles in cellular regulation. In: Grimm B, Porra RJ, Rüdiger W, Scheer H, editors. Biochemistry, biophysics and biological functions of chlorophylls. Dordrecht: Springer; 2006. pp. 223–233.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
