

The Na+/H+ exchanger NHE6 in the endosomal recycling system is involved in the development of apical bile canalicular surface domains in HepG2 cells
- PMID: 20130086
- PMCID: PMC2847532
- DOI: 10.1091/mbc.e09-09-0767
The Na+/H+ exchanger NHE6 in the endosomal recycling system is involved in the development of apical bile canalicular surface domains in HepG2 cells
Abstract
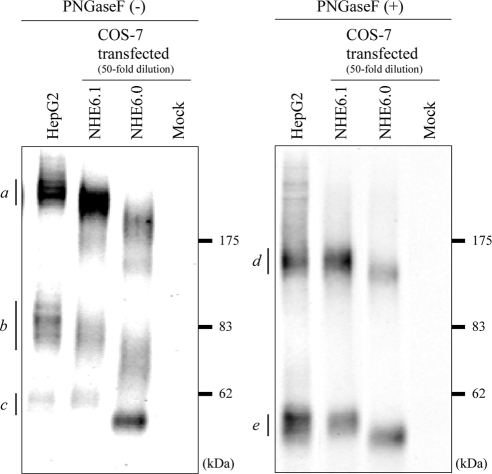
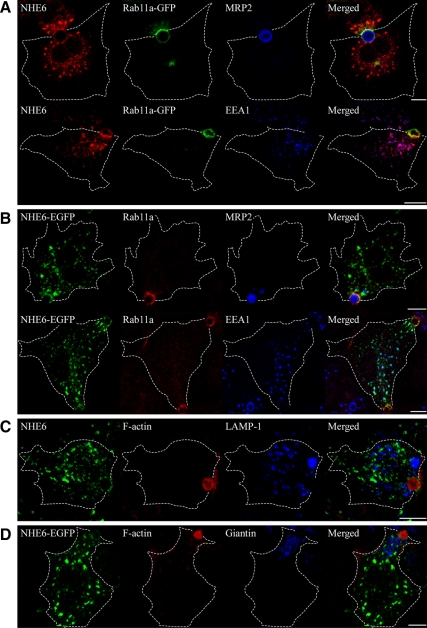
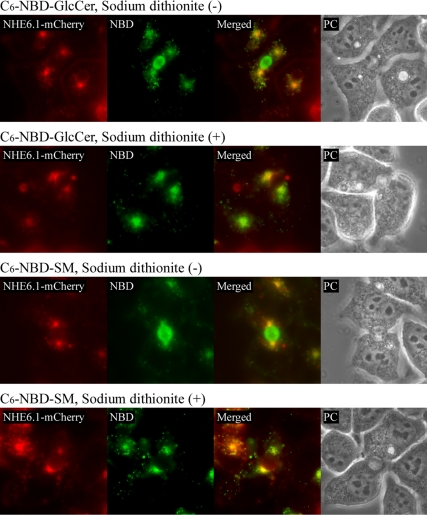
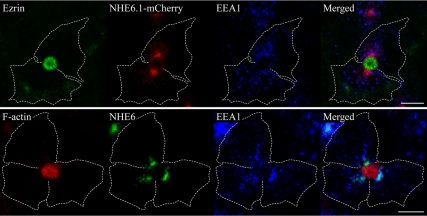
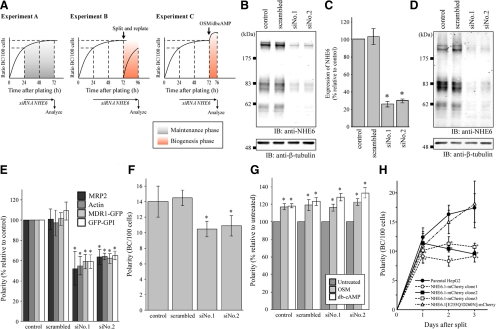
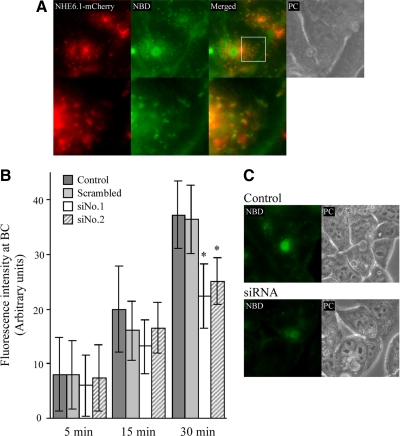
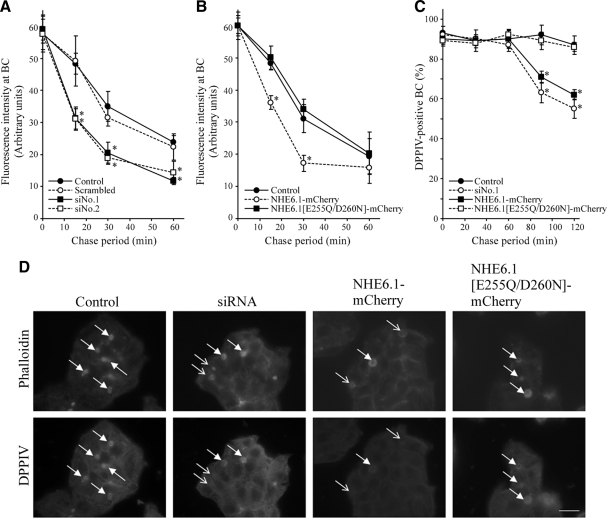
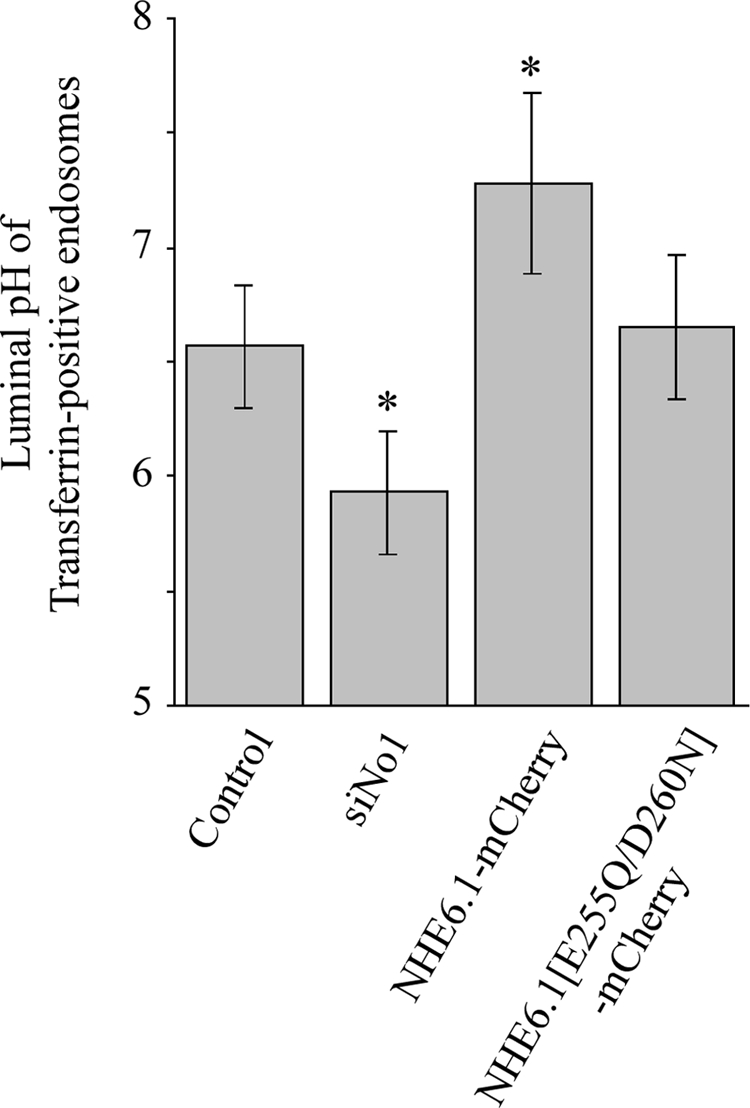
Polarized epithelial cells develop and maintain distinct apical and basolateral surface domains despite a continuous flux of membranes between these domains. The Na(+)/H(+)exchanger NHE6 localizes to endosomes but its function is unknown. Here, we demonstrate that polarized hepatoma HepG2 cells express an NHE6.1 variant that localizes to recycling endosomes and colocalizes with transcytosing bulk membrane lipids. NHE6.1 knockdown or overexpression decreases or increases recycling endosome pH, respectively, and inhibits the maintenance of apical, bile canalicular plasma membranes and, concomitantly, apical lumens. NHE6.1 knockdown or overexpression has little effect on the de novo biogenesis of apical surface domains. NHE6.1 knockdown does not inhibit basolateral-to-apical transcytosis of bulk membrane lipids, but it does promote their progressive loss from the apical surface, leaving cells unable to efficiently retain bulk membrane and bile canalicular proteins at the apical surface. The data suggest that a limited range of endosome pH mediated by NHE6.1 is important for securing the polarized distribution of membrane lipids at the apical surface and maintenance of apical bile canaliculi in HepG2 cells and hence cell polarity. This study underscores the emerging role of the endosomal recycling system in apical surface development and identifies NHE6 as a novel regulatory protein in this process.
Figures
Similar articles
-
Loss of Christianson Syndrome Na+/H+ Exchanger 6 (NHE6) Causes Abnormal Endosome Maturation and Trafficking Underlying Lysosome Dysfunction in Neurons.J Neurosci. 2021 Nov 3;41(44):9235-9256. doi: 10.1523/JNEUROSCI.1244-20.2021. Epub 2021 Sep 15. J Neurosci. 2021. PMID: 34526390 Free PMC article.
-
The Na+/H+ exchanger NHE6 modulates endosomal pH to control processing of amyloid precursor protein in a cell culture model of Alzheimer disease.J Biol Chem. 2015 Feb 27;290(9):5311-27. doi: 10.1074/jbc.M114.602219. Epub 2015 Jan 5. J Biol Chem. 2015. PMID: 25561733 Free PMC article.
-
Na+/H+ exchanger isoform 6 (NHE6/SLC9A6) is involved in clathrin-dependent endocytosis of transferrin.Am J Physiol Cell Physiol. 2011 Dec;301(6):C1431-44. doi: 10.1152/ajpcell.00154.2011. Epub 2011 Aug 31. Am J Physiol Cell Physiol. 2011. PMID: 21881004
-
Emerging links between endosomal pH and cancer.Cancer Metastasis Rev. 2020 Jun;39(2):519-534. doi: 10.1007/s10555-020-09870-1. Cancer Metastasis Rev. 2020. PMID: 32253638 Free PMC article. Review.
-
Recycling endosomes in apical plasma membrane domain formation and epithelial cell polarity.Trends Cell Biol. 2010 Oct;20(10):618-26. doi: 10.1016/j.tcb.2010.08.004. Trends Cell Biol. 2010. PMID: 20833047 Review.
Cited by
-
Dexamethasone disrupts intracellular pH homeostasis to delay coronavirus infectious bronchitis virus cell entry via sodium hydrogen exchanger 3 activation.J Virol. 2025 Jun 17;99(6):e0189424. doi: 10.1128/jvi.01894-24. Epub 2025 May 9. J Virol. 2025. PMID: 40340398 Free PMC article.
-
Endosomal acidification by Na+/H+ exchanger NHE5 regulates TrkA cell-surface targeting and NGF-induced PI3K signaling.Mol Biol Cell. 2013 Nov;24(21):3435-48. doi: 10.1091/mbc.E12-06-0445. Epub 2013 Sep 4. Mol Biol Cell. 2013. PMID: 24006492 Free PMC article.
-
Protons to Patients: targeting endosomal Na+ /H+ exchangers against COVID-19 and other viral diseases.FEBS J. 2021 Sep;288(17):5071-5088. doi: 10.1111/febs.16163. FEBS J. 2021. PMID: 34490733 Free PMC article. Review.
-
GGA1 interacts with the endosomal Na+/H+ exchanger NHE6 governing localization to the endosome compartment.J Biol Chem. 2024 Aug;300(8):107552. doi: 10.1016/j.jbc.2024.107552. Epub 2024 Jul 11. J Biol Chem. 2024. PMID: 39002678 Free PMC article.
-
Altered distribution and localization of organellar Na+/H+ exchangers in postmortem schizophrenia dorsolateral prefrontal cortex.Transl Psychiatry. 2023 Feb 2;13(1):34. doi: 10.1038/s41398-023-02336-2. Transl Psychiatry. 2023. PMID: 36732328 Free PMC article.
References
-
- Babia T., Kok J. W., van der Haar M., Kalicharan R., Hoekstra D. Transport of biosynthetic sphingolipids from Golgi to plasma membrane in HT29 cells: involvement of different carrier vesicle populations. Eur. J. Cell Biol. 1994;63:172–181. - PubMed
-
- Barr V. A., Hubbard A. L. Newly synthesized hepatocyte plasma membrane proteins are transported in transcytotic vesicles in the bile duct-ligated rat. Gastroenterology. 1993;105:554–571. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
