

Interdependence of the peroxisome-targeting receptors in Arabidopsis thaliana: PEX7 facilitates PEX5 accumulation and import of PTS1 cargo into peroxisomes
- PMID: 20130089
- PMCID: PMC2847529
- DOI: 10.1091/mbc.e09-08-0672
Interdependence of the peroxisome-targeting receptors in Arabidopsis thaliana: PEX7 facilitates PEX5 accumulation and import of PTS1 cargo into peroxisomes
Abstract
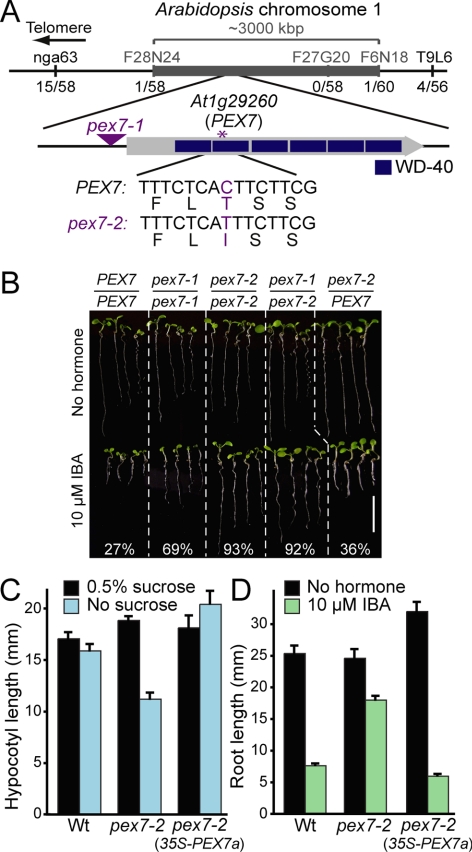
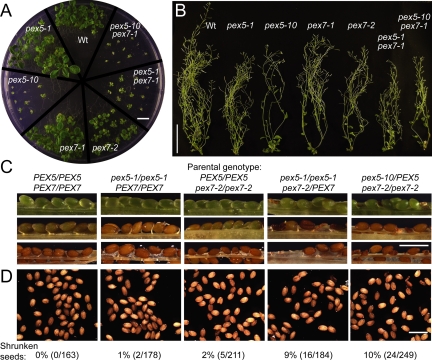
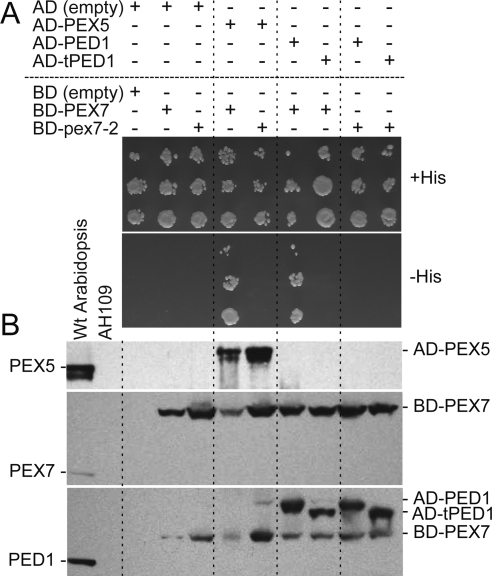
Peroxisomes compartmentalize certain metabolic reactions critical to plant and animal development. The import of proteins from the cytosol into the organelle matrix depends on more than a dozen peroxin (PEX) proteins, with PEX5 and PEX7 serving as receptors that shuttle proteins bearing one of two peroxisome-targeting signals (PTSs) into the organelle. PEX5 is the PTS1 receptor; PEX7 is the PTS2 receptor. In plants and mammals, PEX7 depends on PEX5 binding to deliver PTS2 cargo into the peroxisome. In this study, we characterized a pex7 missense mutation, pex7-2, that disrupts both PEX7 cargo binding and PEX7-PEX5 interactions in yeast, as well as PEX7 protein accumulation in plants. We examined localization of peroxisomally targeted green fluorescent protein derivatives in light-grown pex7 mutants and observed not only the expected defects in PTS2 protein import but also defects in PTS1 import. These PTS1 import defects were accompanied by reduced PEX5 accumulation in light-grown pex7 seedlings. Our data suggest that PEX5 and PTS1 import depend on the PTS2 receptor PEX7 in Arabidopsis and that the environment may influence this dependence. These data advance our understanding of the biogenesis of these essential organelles and provide a possible rationale for the retention of the PTS2 pathway in some organisms.
Figures



References
-
- Adham A. R., Zolman B. K., Millius A., Bartel B. Mutations in Arabidopsis acyl-CoA oxidase genes reveal distinct and overlapping roles in β-oxidation. Plant J. 2005;41:859–874. - PubMed
-
- Asamizu E., Nakamura Y., Sato S., Tabata S. A large scale analysis of cDNA in Arabidopsis thaliana: generation of 12,028 non-redundant expressed sequence tags from normalized and size-selected cDNA libraries. DNA Res. 2000;7:175–180. - PubMed
-
- Ausubel F., Brent R., Kingston R. E., Moore D. D., Seidman J. G., Smith J. A., Struhl K. Current Protocols in Molecular Biology. New York: Greene Publishing Associates and Wiley-Interscience; 1999.
-
- Braverman N., Dodt G., Gould S. J., Valle D. An isoform of Pex5p, the human PTS1 receptor, is required for the import of PTS2 proteins into peroxisomes. Hum. Mol. Genet. 1998;7:1195–1205. - PubMed
-
- Brocard C., Kragler F., Simon M. M., Schuster T., Hartig A. The tetratricopeptide repeat-domain of the PAS10 protein of Saccharomyces cerevisiae is essential for binding the peroxisomal targeting signal -SKL. Biochem. Biophys. Res. Commun. 1994;204:1016–1022. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
