

Genetic fate mapping reveals that the caudal ganglionic eminence produces a large and diverse population of superficial cortical interneurons
- PMID: 20130169
- PMCID: PMC2826846
- DOI: 10.1523/JNEUROSCI.4515-09.2010
Genetic fate mapping reveals that the caudal ganglionic eminence produces a large and diverse population of superficial cortical interneurons
Abstract
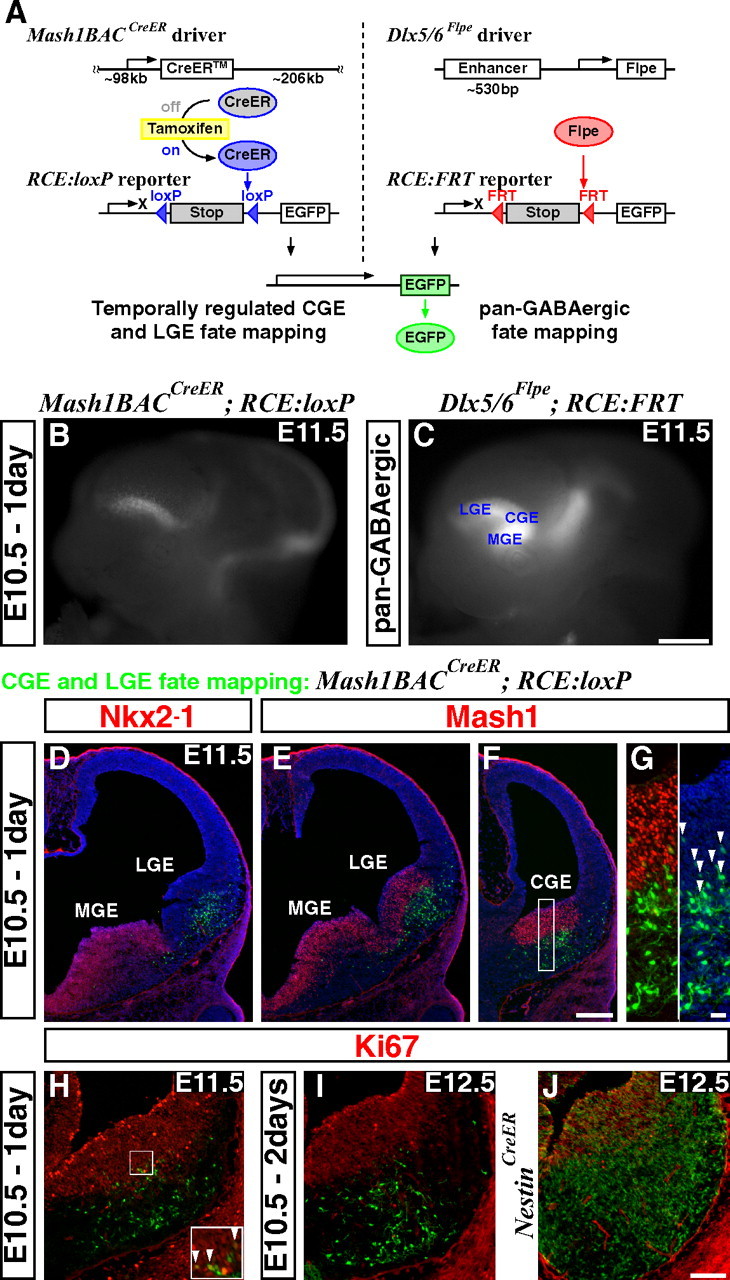
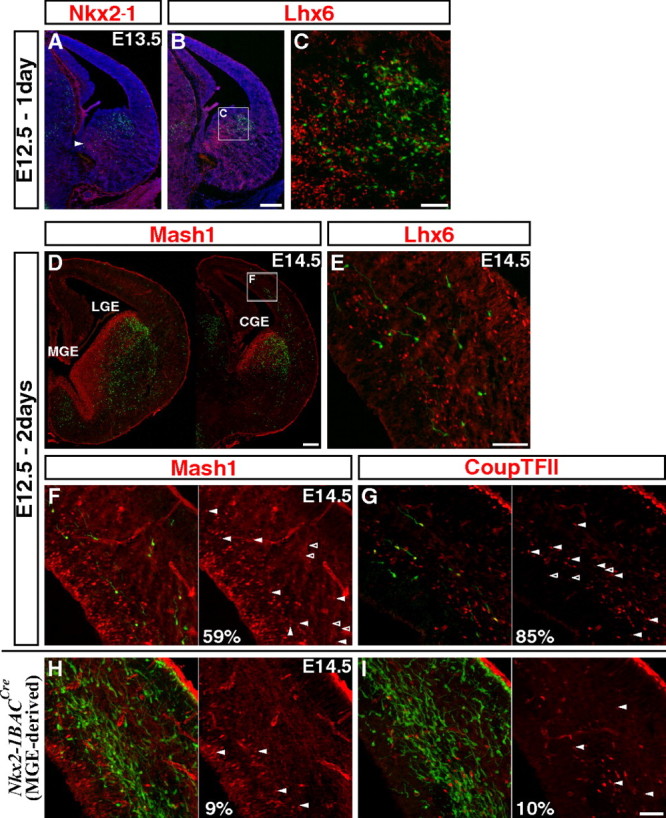
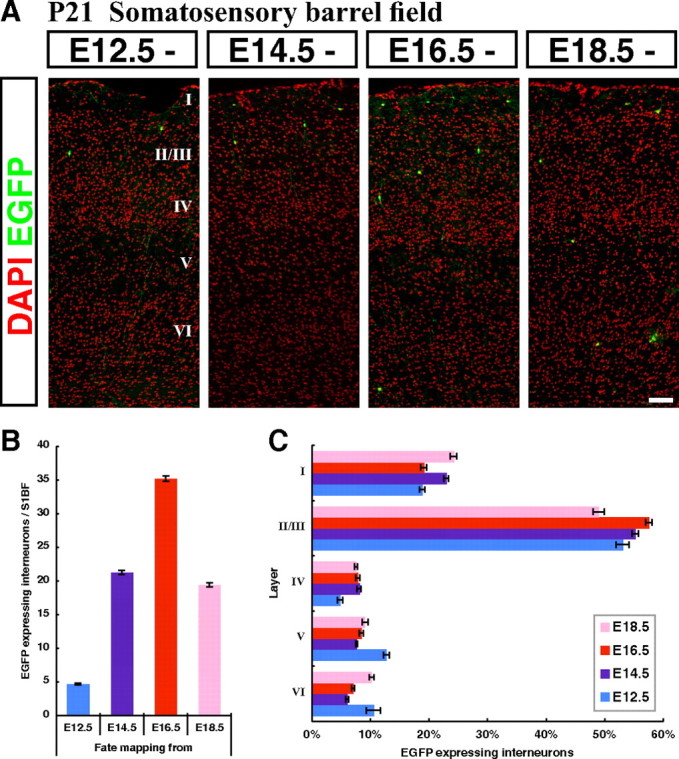
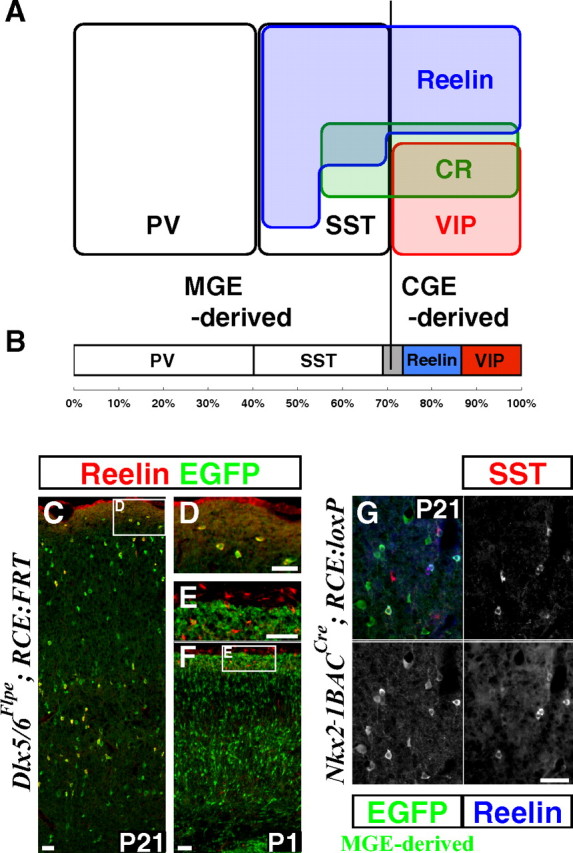
By combining an inducible genetic fate mapping strategy with electrophysiological analysis, we have systematically characterized the populations of cortical GABAergic interneurons that originate from the caudal ganglionic eminence (CGE). Interestingly, compared with medial ganglionic eminence (MGE)-derived cortical interneuron populations, the initiation [embryonic day 12.5 (E12.5)] and peak production (E16.5) of interneurons from this embryonic structure occurs 3 d later in development. Moreover, unlike either pyramidal cells or MGE-derived cortical interneurons, CGE-derived interneurons do not integrate into the cortex in an inside-out manner but preferentially (75%) occupy superficial cortical layers independent of birthdate. In contrast to previous estimates, CGE-derived interneurons are both considerably greater in number (approximately 30% of all cortical interneurons) and diversity (comprised by at least nine distinct subtypes). Furthermore, we found that a large proportion of CGE-derived interneurons, including the neurogliaform subtype, express the glycoprotein Reelin. In fact, most CGE-derived cortical interneurons express either Reelin or vasoactive intestinal polypeptide. Thus, in conjunction with previous studies, we have now determined the spatial and temporal origins of the vast majority of cortical interneuron subtypes.
Figures


Comment in
-
Last but not least: cortical interneurons from caudal ganglionic eminence.J Neurosci. 2010 Jun 2;30(22):7449-50. doi: 10.1523/JNEUROSCI.1677-10.2010. J Neurosci. 2010. PMID: 20519518 Free PMC article. No abstract available.
References
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Angevine JB, II, Sidman RL. Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse. Nature. 1961;192:766–768. - PubMed
-
- Ascoli GA, Alonso-Nanclares L, Anderson SA, Barrionuevo G, Benavides-Piccione R, Burkhalter A, Buzsáki G, Cauli B, Defelipe J, Fairén A, Feldmeyer D, Fishell G, Fregnac Y, Freund TF, Gardner D, Gardner EP, Goldberg JH, Helmstaedter M, Hestrin S, Karube F, et al. Petilla terminology: nomenclature of features of GABAergic interneurons of the cerebral cortex. Nat Rev Neurosci. 2008;9:557–568. - PMC - PubMed
-
- Bai CB, Auerbach W, Lee JS, Stephen D, Joyner AL. Gli2, but not Gli1, is required for initial Shh signaling and ectopic activation of the Shh pathway. Development. 2002;129:4753–4761. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases