

Dopamine and memory: modulation of the persistence of memory for novel hippocampal NMDA receptor-dependent paired associates
- PMID: 20130171
- PMCID: PMC6633999
- DOI: 10.1523/JNEUROSCI.2721-09.2010
Dopamine and memory: modulation of the persistence of memory for novel hippocampal NMDA receptor-dependent paired associates
Abstract
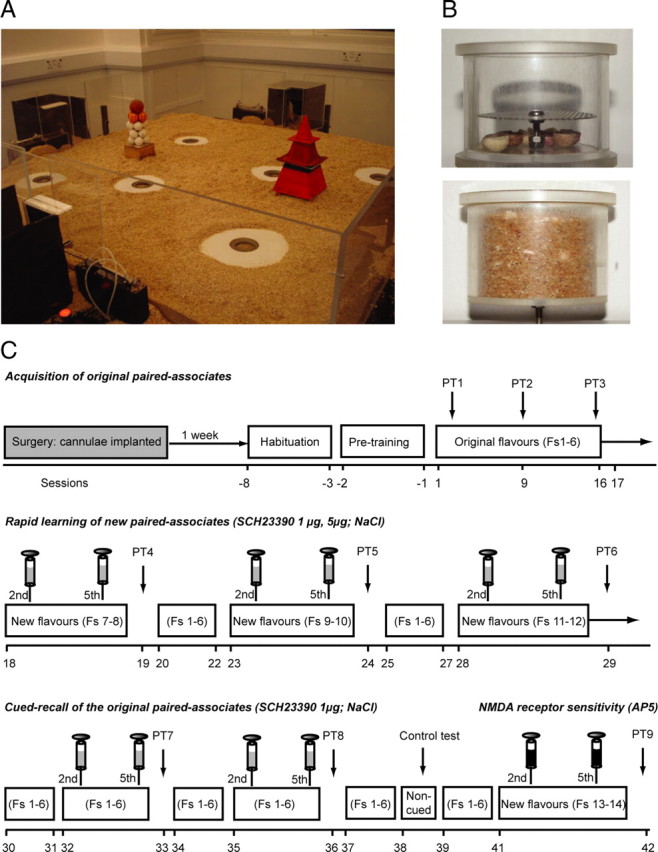
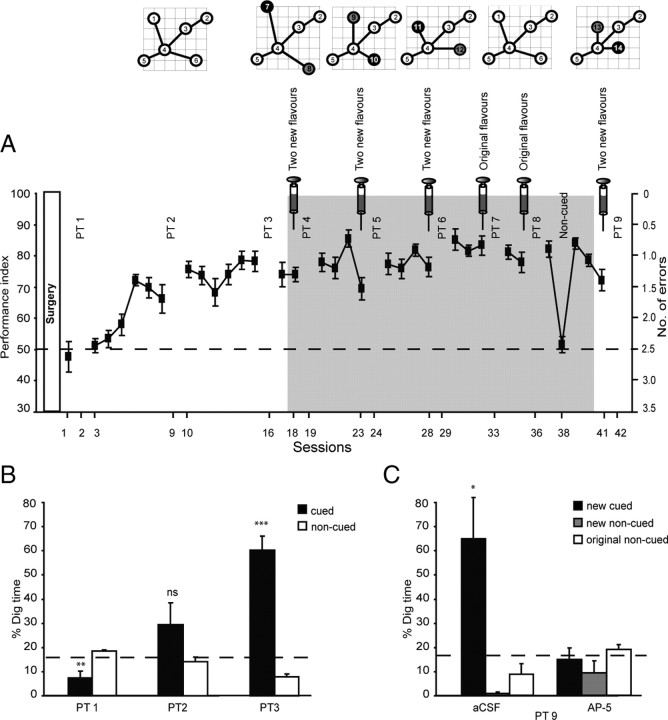
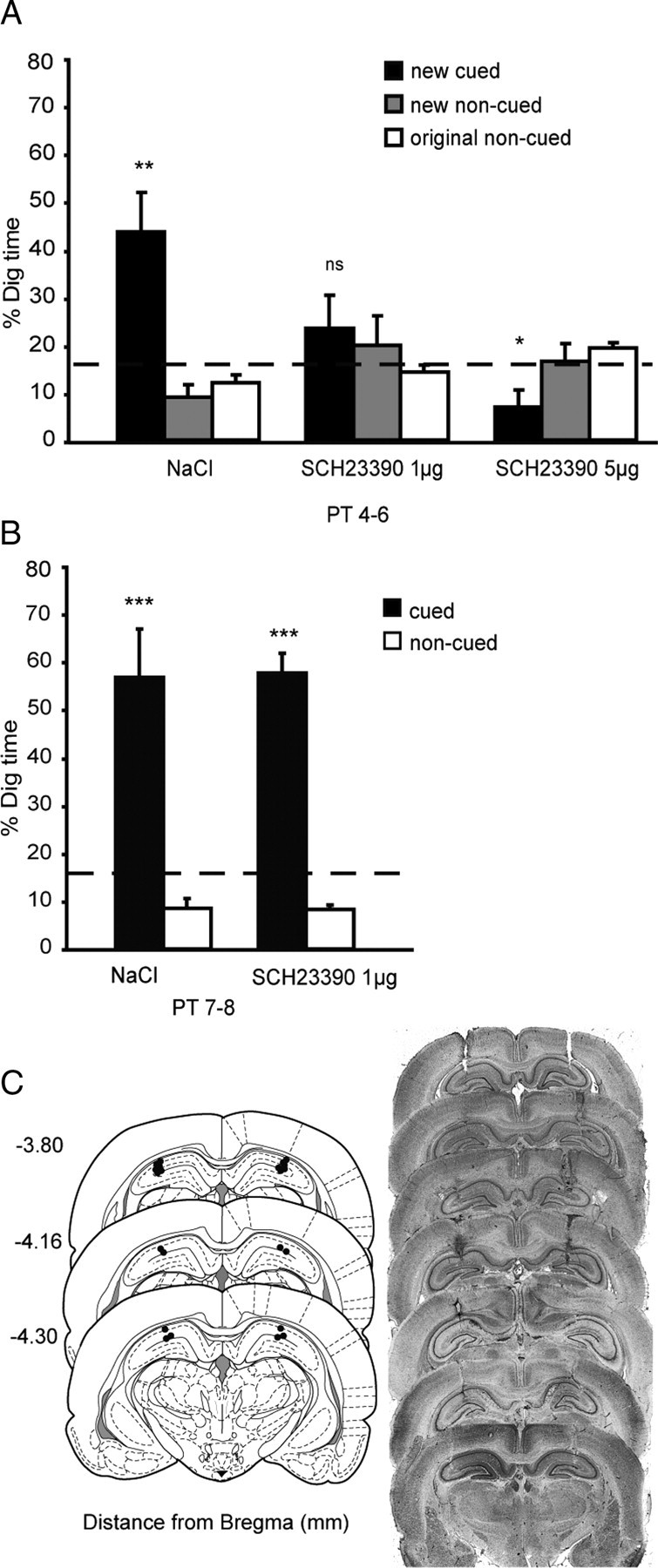
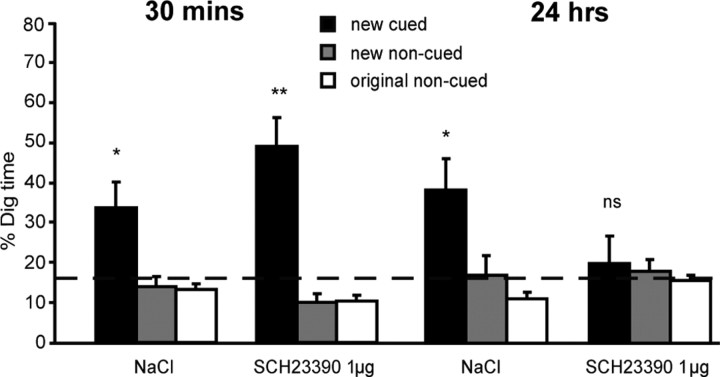
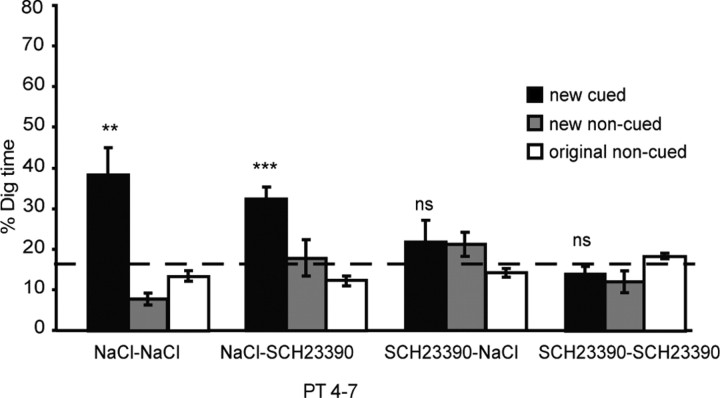
Three experiments investigated the role in memory processing of dopamine (DA) afferents to the hippocampus (HPC) that arise from the ventral tegmental area. One hypothesis is that D(1)/D(5) receptor activation in HPC is necessary for the encoding of novel, episodic-like information; the other is that DA activation ensures the greater temporal persistence of transient hippocampal memory traces. Rats (n = 35) were trained, in separate experiments using an episodic-like memory task, to learn six paired associates (PAs) in an "event arena" involving a repeated association between specific flavors of food and locations in space. After 6 weeks of training, rats had learned a "schema" such that two new paired associates could be acquired in a single trial in one session (episodic-like memory). We show that encoding of novel PAs is sensitive to intrahippocampal microinfusion of the NMDA antagonist d-AP-5. Experiment 1 established that intrahippocampal infusion of the D(1)/D(5) dopaminergic antagonist SCH23390 [R(+)-7-chloro-8-hydroxy-3-methyl-1-phenyl-2,3,4,5-tetrahydro-1H-3-benzazepine hydrochloride] before encoding of new PAs caused impaired memory 24 h later but that SCH23390 had no effect on the later memory of previously established PAs. Experiment 2 established that SCH23390 modulated the persistence of new memories over time (30 min vs 24 h) rather than affecting initial encoding. Experiment 3 revealed that the impact of SCH23390 was not mediated by state dependence nor had an effect on memory retrieval. These findings support the second hypothesis and establish that persistent, long-term memory of rapid, hippocampal-mediated acquisition of new paired associates requires activation of D(1)/D(5) receptors in HPC at or around the time of encoding.
Figures
References
-
- Adcock RA, Thangavel A, Whitfield-Gabrieli S, Knutson B, Gabrieli JD. Reward-motivated learning: mesolimbic activation precedes memory formation. Neuron. 2006;50:507–517. - PubMed
-
- Bernabeu R, Bevilaqua L, Ardenghi P, Bromberg E, Schmitz P, Bianchin M, Izquierdo I, Medina JH. Involvement of hippocampal cAMP/cAMP-dependent protein kinase signaling pathways in a late memory consolidation phase of aversively motivated learning in rats. Proc Natl Acad Sci U S A. 1997;94:7041–7046. - PMC - PubMed
-
- Bliss TV, Collingridge GL. A synaptic model of memory: long-term potentiation in the hippocampus. Nature. 1993;361:31–39. - PubMed
-
- Bontempi B, Laurent-Demir C, Destrade C, Jaffard R. Time-dependent reorganization of brain circuitry underlying long-term memory storage. Nature. 1999;400:671–675. - PubMed
-
- Buhot MC, Martin S, Segu L. Role of serotonin in memory impairment. Ann Med. 2000;32:210–221. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical