

MicroRNA regulation of IFN-beta protein expression: rapid and sensitive modulation of the innate immune response
- PMID: 20130213
- PMCID: PMC3076721
- DOI: 10.4049/jimmunol.0902712
MicroRNA regulation of IFN-beta protein expression: rapid and sensitive modulation of the innate immune response
Abstract
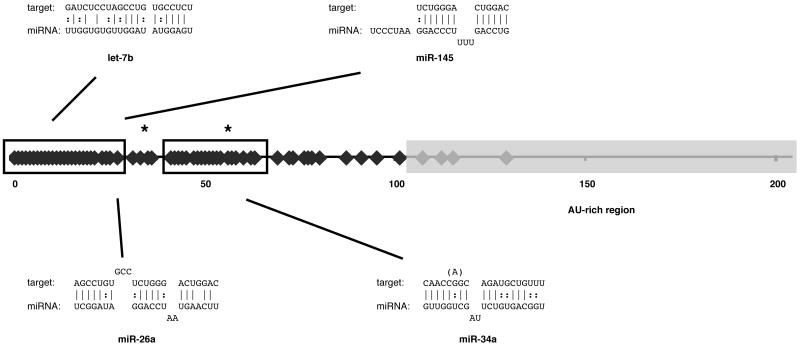
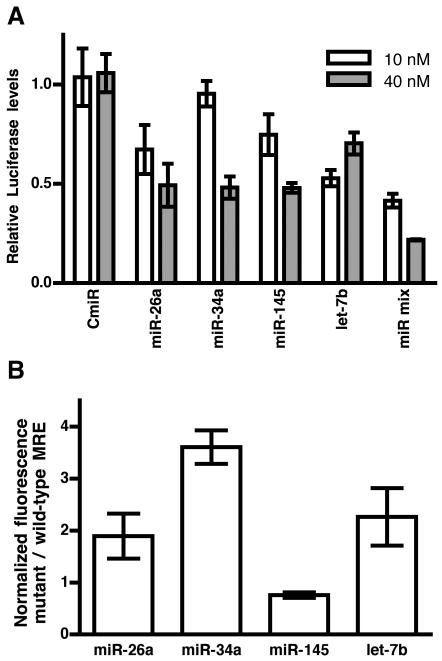
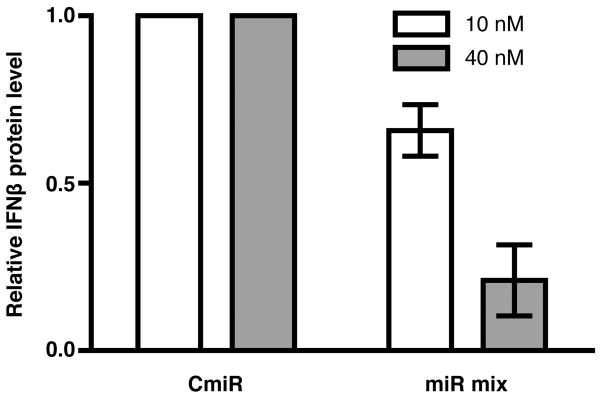
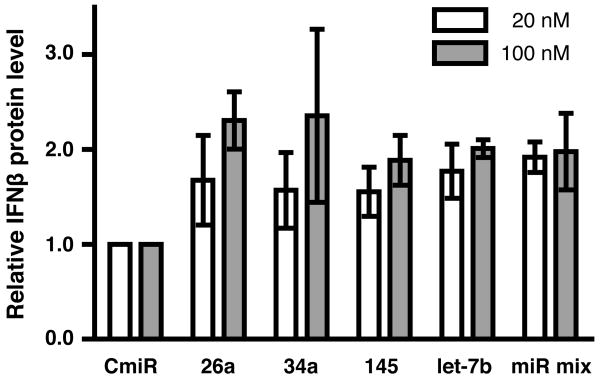
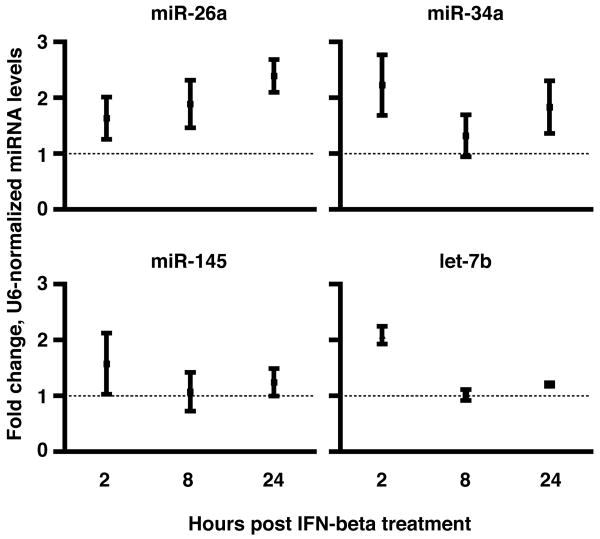
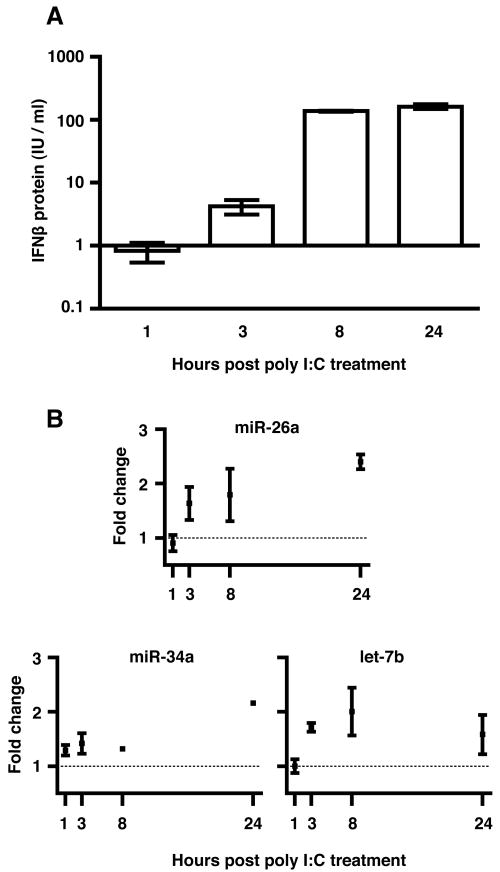
IFN-beta production is an inaugural event in the innate immune response to viral infections, with relatively small fold changes in IFN-beta expression resulting in the activation of important antiviral signaling cascades. In our rapid SIV/macaque model of HIV encephalitis, the virus enters the CNS within 4 d of infection, accompanied by a marked IFN-beta response that wanes as SIV replication is controlled. The centrality of IFN-beta to the innate antiviral response in the CNS combines with the potential inflammatory damage associated with long-term activation of this pathway to suggest that IFN-beta may be subject to regulatory fine-tuning in addition to well-established transcriptional and message stability mechanisms of regulation. In this paper, we present for the first time evidence that microRNAs (miRNAs), including miR-26a, -34a, -145, and let-7b, may directly regulate IFN-beta in human and macaque cells. In primary primate macrophages, the main cell type implicated in HIV and SIV infection in the CNS, specific miRNAs reduce, whereas miRNA inhibitors enhance, IFN-beta protein production. The potential biologic significance of this regulation is supported by evidence of an apparent negative feedback loop, with increased expression of three IFN-beta-regulating miRNAs by primate macrophages exposed to recombinant IFN-beta or stimulated to produce IFN-beta. Thus, miRNAs may contribute significantly to the regulation of IFN-beta in innate immune responses.
Figures
Similar articles
-
Suppressor of cytokine signaling 3 inhibits antiviral IFN-beta signaling to enhance HIV-1 replication in macrophages.J Immunol. 2010 Aug 15;185(4):2393-404. doi: 10.4049/jimmunol.0903563. Epub 2010 Jul 14. J Immunol. 2010. PMID: 20631305 Free PMC article.
-
SIV replication is directly downregulated by four antiviral miRNAs.Retrovirology. 2013 Aug 29;10:95. doi: 10.1186/1742-4690-10-95. Retrovirology. 2013. PMID: 23988154 Free PMC article.
-
Yin Yang 1 Dynamically Regulates Antiviral Innate Immune Responses During Viral Infection.Cell Physiol Biochem. 2017;44(2):607-617. doi: 10.1159/000485116. Epub 2017 Nov 20. Cell Physiol Biochem. 2017. PMID: 29161701
-
microRNA control of interferons and interferon induced anti-viral activity.Mol Immunol. 2013 Dec;56(4):781-93. doi: 10.1016/j.molimm.2013.07.009. Epub 2013 Aug 23. Mol Immunol. 2013. PMID: 23962477 Review.
-
Regulation of the MIR155 host gene in physiological and pathological processes.Gene. 2013 Dec 10;532(1):1-12. doi: 10.1016/j.gene.2012.12.009. Epub 2012 Dec 14. Gene. 2013. PMID: 23246696 Review.
Cited by
-
miRNA profiles of monocyte-lineage cells are consistent with complicated roles in HIV-1 restriction.Viruses. 2012 Sep 25;4(10):1844-64. doi: 10.3390/v4101844. Viruses. 2012. PMID: 23202444 Free PMC article.
-
A major locus on chromosome 3p22 conferring predisposition to human herpesvirus 8 infection.Eur J Hum Genet. 2012 Jun;20(6):690-5. doi: 10.1038/ejhg.2011.260. Epub 2012 Jan 18. Eur J Hum Genet. 2012. PMID: 22258534 Free PMC article.
-
Liposome delivery of microRNA-145 to mesenchymal stem cells leads to immunological off-target effects mediated by RIG-I.Mol Ther. 2013 Jun;21(6):1169-81. doi: 10.1038/mt.2013.55. Epub 2013 Apr 9. Mol Ther. 2013. PMID: 23568258 Free PMC article.
-
Analysis of microRNA and Gene Expression Profiles in Multiple Sclerosis: Integrating Interaction Data to Uncover Regulatory Mechanisms.Sci Rep. 2016 Oct 3;6:34512. doi: 10.1038/srep34512. Sci Rep. 2016. PMID: 27694855 Free PMC article.
-
Differential microRNA expression in the prefrontal cortex of mouse offspring induced by glyphosate exposure during pregnancy and lactation.Exp Ther Med. 2018 Mar;15(3):2457-2467. doi: 10.3892/etm.2017.5669. Epub 2017 Dec 21. Exp Ther Med. 2018. PMID: 29467848 Free PMC article.
References
-
- Stacey AR, Norris PJ, Qin L, Haygreen EA, Taylor E, Heitman J, Lebedeva M, DeCamp A, Li D, Grove D, Self SG, Borrow P. Induction of a striking systemic cytokine cascade prior to peak viremia in acute human immunodeficiency virus type 1 infection, in contrast to more modest and delayed responses in acute hepatitis B and C virus infections. J Virol. 2009;83:3719–3733. - PMC - PubMed
-
- Barber SA, Herbst DS, Bullock BT, Gama L, Clements JE. Innate immune responses and control of acute simian immunodeficiency virus replication in the central nervous system. J Neurovirol. 2004;10 1:15–20. - PubMed
-
- Barber SA, Gama L, Li M, Voelker T, Anderson JE, Zink MC, Tarwater PM, Carruth LM, Clements JE. Longitudinal analysis of simian immunodeficiency virus (SIV) replication in the lungs: compartmentalized regulation of SIV. J Infect Dis. 2006;194:931–938. - PubMed
-
- Honda Y, Rogers L, Nakata K, Zhao BY, Pine R, Nakai Y, Kurosu K, Rom WN, Weiden M. Type I interferon induces inhibitory 16-kD CCAAT/enhancer binding protein (C/EBP)beta, repressing the HIV-1 long terminal repeat in macrophages: pulmonary tuberculosis alters C/EBP expression, enhancing HIV-1 replication. J Exp Med. 1998;188:1255–1265. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
