

Absence of estrogen receptor alpha leads to physiological alterations in the mouse epididymis and consequent defects in sperm function
- PMID: 20130267
- PMCID: PMC2857635
- DOI: 10.1095/biolreprod.109.079889
Absence of estrogen receptor alpha leads to physiological alterations in the mouse epididymis and consequent defects in sperm function
Abstract
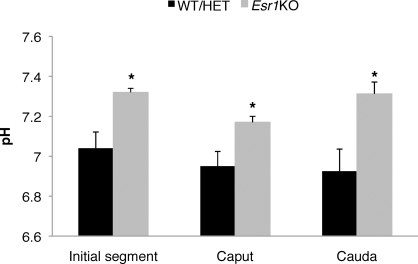
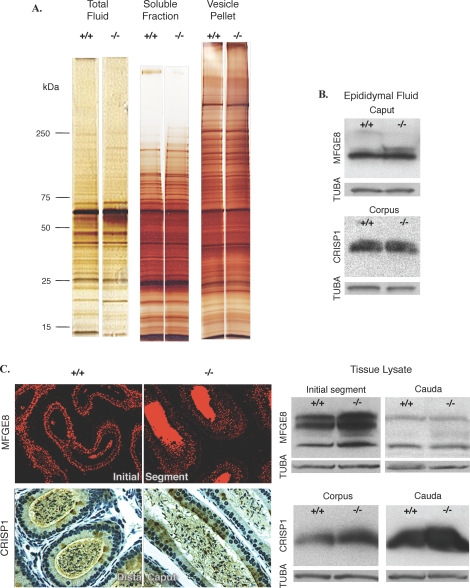
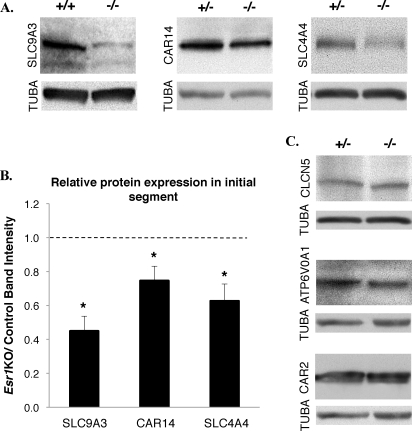
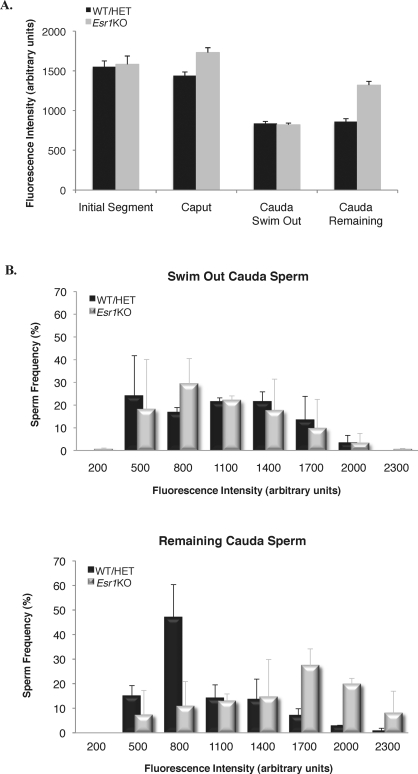
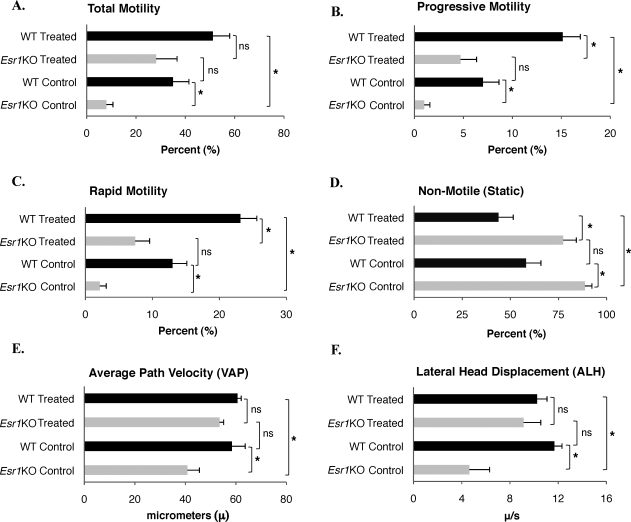
Male mice deficient in ESR1 (ERalpha) (Esr1KO mice) are infertile, and sperm recovered from the cauda epididymis exhibit reduced motility and fail to fertilize eggs in vitro. These effects on sperm appear to result from defective epididymal function and not a direct effect on spermatogenesis, as Esr1KO germ cells transplanted into wild-type testes yield normal offspring. We hypothesized that the previously described defect in efferent duct fluid reabsorption would lead to alterations in the epididymal fluid milieu, which would negatively impact sperm function. Analysis of the epididymal fluid revealed that the Esr1KO maintains a higher luminal pH throughout the epididymis, confirming an inability of the efferent ducts and/or epididymis to properly acidify the luminal contents. Subsequent studies showed that these abnormalities were not the result of global defects in epididymal function since protein secretion by the Esr1KO epididymis appeared normal as judged by SDS-PAGE of total secreted proteins and by immunoblotting of candidate secreted proteins. To gain insight into the basis of the aberrant fluid homeostasis in the Esr1KO epididymis, the expression of several enzymes and transporters known to be involved in acid/base regulation were analyzed. The levels of SLC9A3 (NHE3) as well as carbonic anhydrase XIV and SLC4A4 (NBC1) were all reduced in the proximal portion of the Esr1KO epididymis, while other components appeared unaffected, including other ion transporters and ATP6V0A1 (V-ATPase). The altered luminal milieu of the Esr1KO epididymis was shown to lead to a corresponding increase in the intracellular pH of Esr1KO sperm, relative to sperm from control animals. Since pH and bicarbonate ions are critical regulators of sperm cAMP levels and motility, we attempted to bypass the abnormal luminal and intracellular environment by supplementing sperm with exogenous cAMP. This treatment rescued all defective motility parameters, as assayed by CASA, further showing that motility defects are not intrinsic to the sperm but, rather, result from the abnormal epididymal milieu.
Figures
References
-
- Eddy EM, Washburn TF, Bunch DO, Goulding EH, Gladen BC, Lubahn DB, Korach KS.Targeted disruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis and infertility. Endocrinology 1996; 137: 4796–4805. - PubMed
-
- Mahato D, Goulding EH, Korach KS, Eddy EM.Estrogen receptor-alpha is required by the supporting somatic cells for spermatogenesis. Mol Cell Endocrinol 2001; 178: 57–63. - PubMed
-
- Mahato D, Goulding EH, Korach KS, Eddy EM.Spermatogenic cells do not require estrogen receptor-alpha for development or function. Endocrinology 2000; 141: 1273–1276. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
