

Rapid elimination of the histone variant MacroH2A from somatic cell heterochromatin after nuclear transfer
- PMID: 20132012
- PMCID: PMC2998967
- DOI: 10.1089/cell.2009.0043
Rapid elimination of the histone variant MacroH2A from somatic cell heterochromatin after nuclear transfer
Abstract
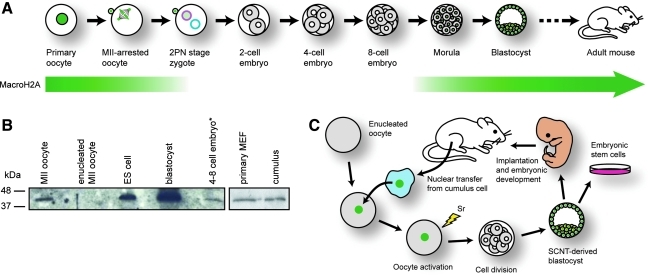
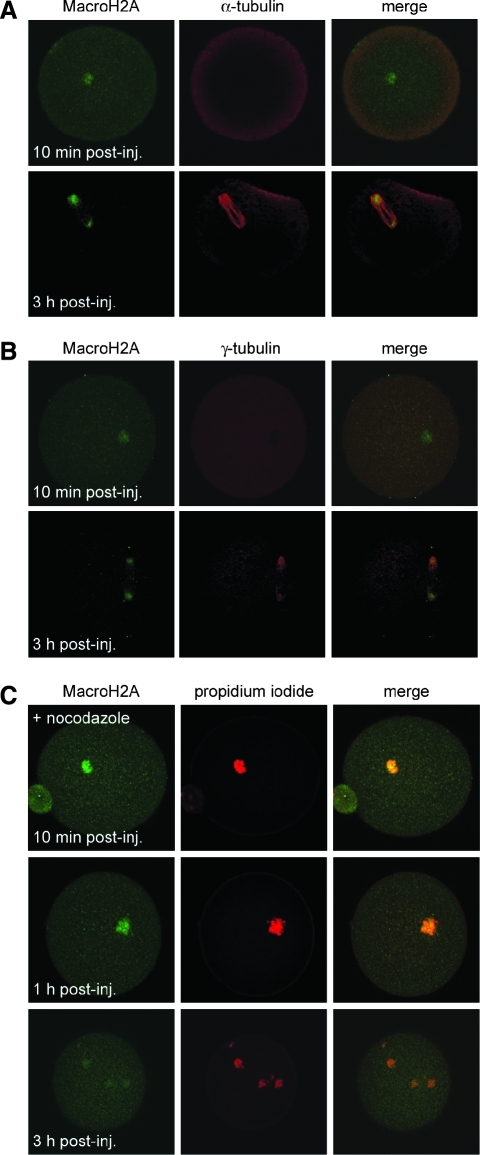
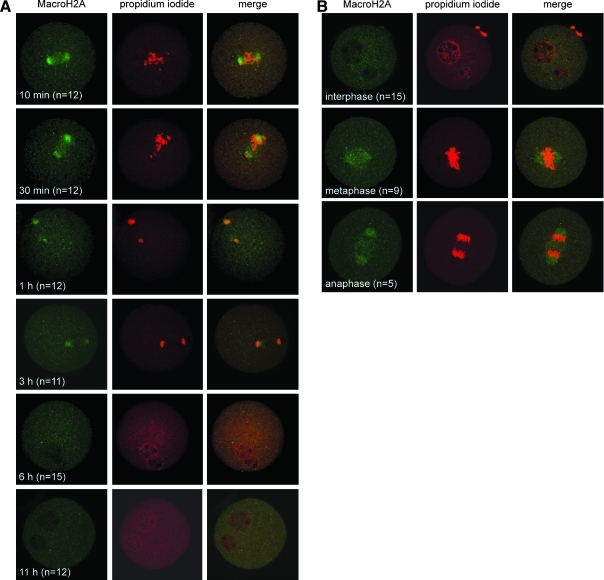
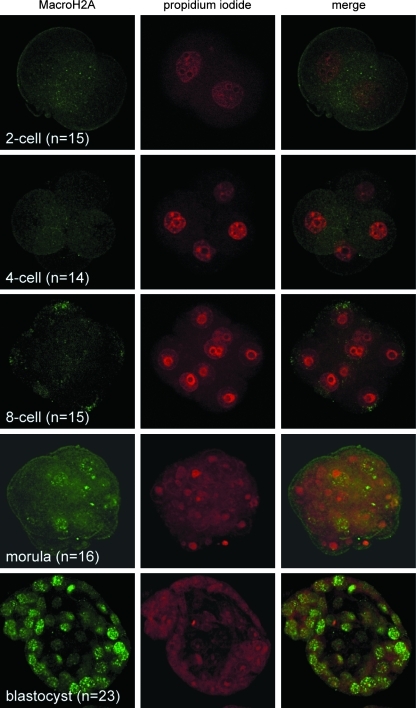
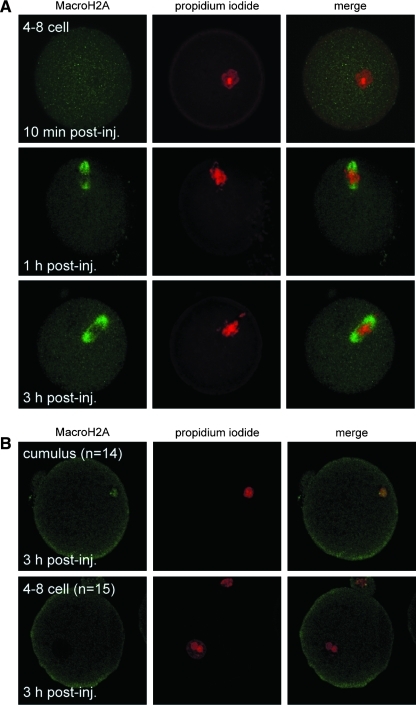
Oocytes contain a maternal store of the histone variant MacroH2A, which is eliminated from zygotes shortly after fertilization. Preimplantation embryos then execute three cell divisions without MacroH2A before the onset of embryonic MacroH2A expression at the 16-cell stage. During subsequent development, MacroH2A is expressed in most cells, where it is assembled into facultative heterochromatin. Because differentiated cells contain heterochromatin rich in MacroH2A, we investigated the fate of MacroH2A during somatic cell nuclear transfer (SCNT). The results show that MacroH2A is rapidly eliminated from the chromosomes of transplanted somatic cell nuclei by a process in which MacroH2A is first stripped from chromosomes, and then degraded. Furthermore, MacroH2A is eliminated from transplanted nuclei by a mechanism requiring intact microtubules and nuclear envelope break down. Preimplantation SCNT embryos express endogenous MacroH2A once they reach the morula stage, similar to the timing observed in embryos produced by natural fertilization. We also show that the ability to reprogram somatic cell heterochromatin by SCNT is tied to the developmental stage of recipient cell cytoplasm because enucleated zygotes fail to support depletion of MacroH2A from transplanted somatic nuclei. Together, the results indicate that nuclear reprogramming by SCNT utilizes the same chromatin remodeling mechanisms that act upon the genome immediately after fertilization.
Figures

References
-
- Angelov D. Molla A. Perche P.Y., et al. The histone variant macroH2A interferes with transcription factor binding and SWI/SNF nucleosome remodeling. Mol. Cell. 2003;11:1033–1041. - PubMed
-
- Armstrong L. Lako M. Dean W., et al. Epigenetic modification is central to genome reprogramming in somatic cell nuclear transfer. Stem Cells. 2006;24:805–814. - PubMed
-
- Byrne J.A. Simonsson S. Western P.S., et al. Nuclei of adult mammalian somatic cells are directly reprogrammed to oct-4 stem cell gene expression by amphibian oocytes. Curr. Biol. 2003;13:1206–1213. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
