

The structure of innate vocalizations in Foxp2-deficient mouse pups
- PMID: 20132318
- PMCID: PMC2895353
- DOI: 10.1111/j.1601-183X.2010.00570.x
The structure of innate vocalizations in Foxp2-deficient mouse pups
Abstract
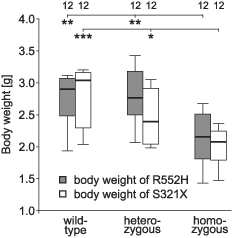
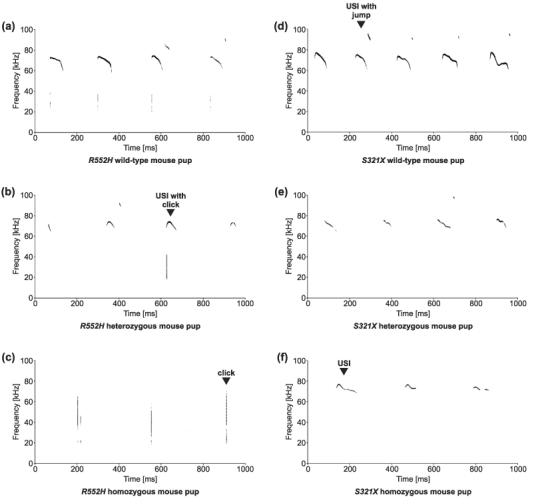
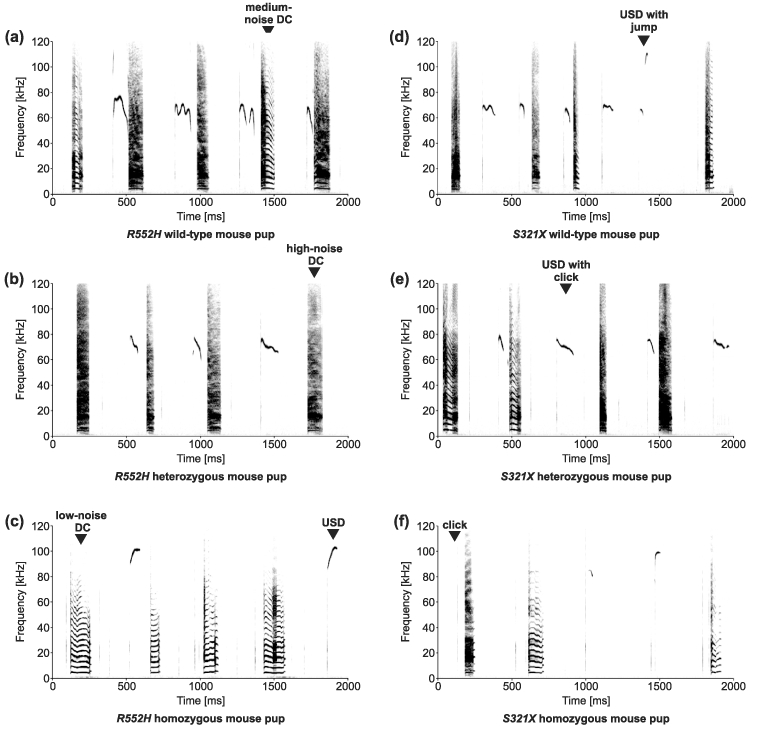
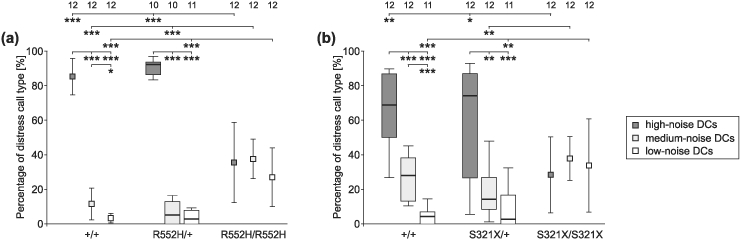
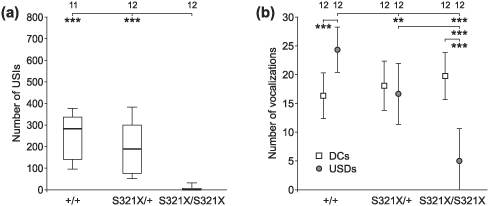
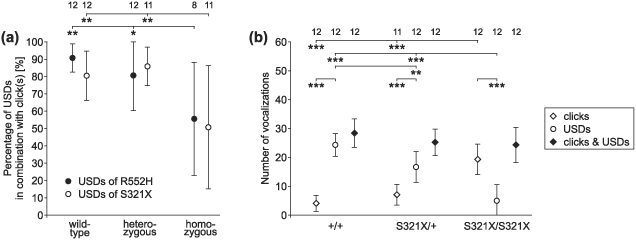
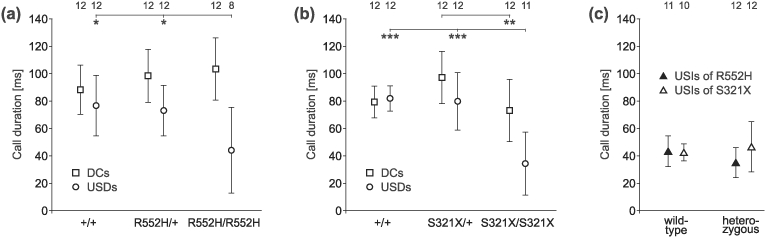
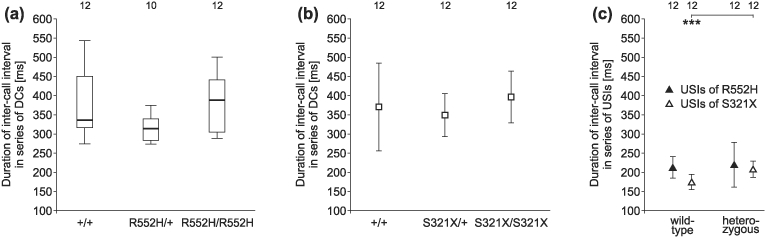
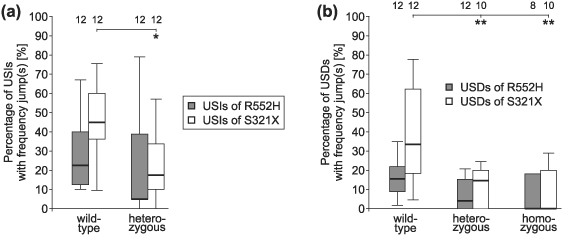
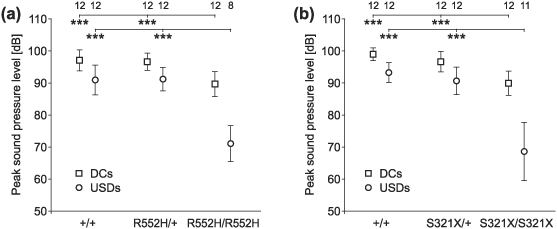
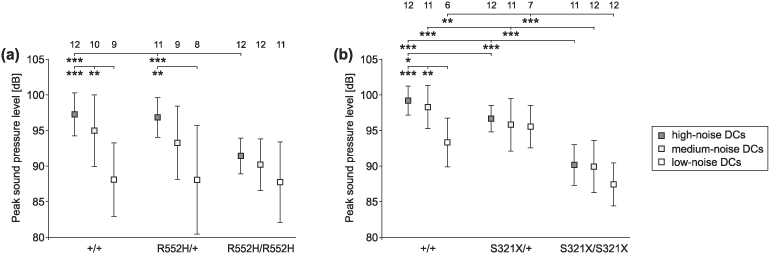
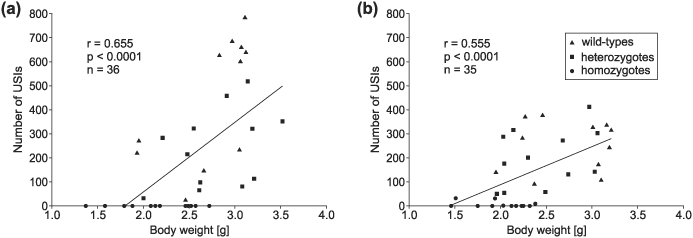
Heterozygous mutations of the human FOXP2 gene are implicated in a severe speech and language disorder. Aetiological mutations of murine Foxp2 yield abnormal synaptic plasticity and impaired motor-skill learning in mutant mice, while knockdown of the avian orthologue in songbirds interferes with auditory-guided vocal learning. Here, we investigate influences of two distinct Foxp2 point mutations on vocalizations of 4-day-old mouse pups (Mus musculus). The R552H missense mutation is identical to that causing speech and language deficits in a large well-studied human family, while the S321X nonsense mutation represents a null allele that does not produce Foxp2 protein. We ask whether vocalizations, based solely on innate mechanisms of production, are affected by these alternative Foxp2 mutations. Sound recordings were taken in two different situations: isolation and distress, eliciting a range of call types, including broadband vocalizations of varying noise content, ultrasonic whistles and clicks. Sound production rates and several acoustic parameters showed that, despite absence of functional Foxp2, homozygous mutants could vocalize all types of sounds in a normal temporal pattern, but only at comparably low intensities. We suggest that altered vocal output of these homozygotes may be secondary to developmental delays and somatic weakness. Heterozygous mutants did not differ from wild-types in any of the measures that we studied (R552H ) or in only a few (S321X ), which were in the range of differences routinely observed for different mouse strains. Thus, Foxp2 is not essential for the innate production of emotional vocalizations with largely normal acoustic properties by mouse pups.
Figures
References
-
- Bell RW, Nitschke W, Zachman T. Ultrasounds in three inbred strains of young mice. Behav Biol. 1972;7:805–814. - PubMed
-
- Branchi I, Santucci D, Puopolo M, Alleva E. Neonatal behaviours associated with ultrasonic vocalizations in mice (Mus musculus): a slow-motion analysis. Dev Psychobiol. 2004;44:37–44. - PubMed
-
- Brunelli SA. Selective breeding for an infant phenotype: rat pup ultrasonic vocalization (USV) Behav Genet. 2005;35:53–65. - PubMed
-
- D’Udine B, Robinson DJ, Oliverio A. An analysis of single-gene effects on audible and ultrasonic vocalizations in the mouse. Behav Neural Biol. 1982;36:197–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
