

Positioned and G/C-capped poly(dA:dT) tracts associate with the centers of nucleosome-free regions in yeast promoters
- PMID: 20133331
- PMCID: PMC2847750
- DOI: 10.1101/gr.103226.109
Positioned and G/C-capped poly(dA:dT) tracts associate with the centers of nucleosome-free regions in yeast promoters
Abstract
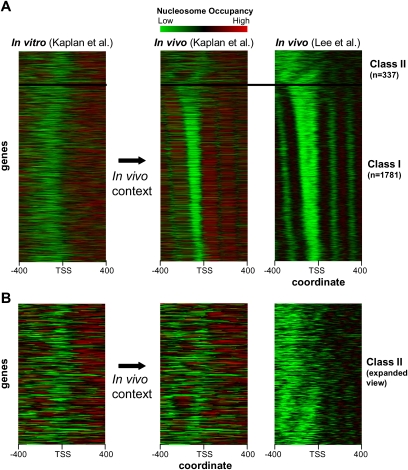
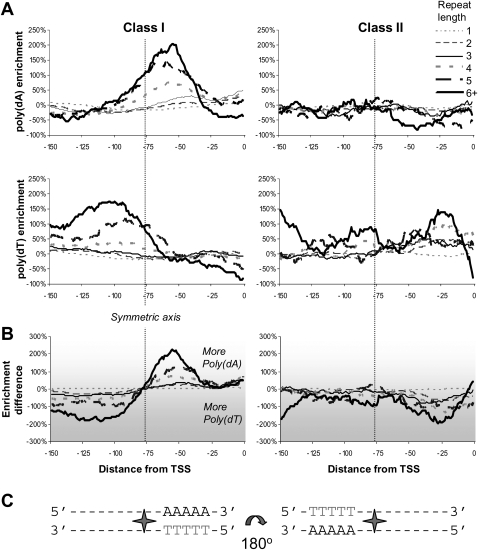
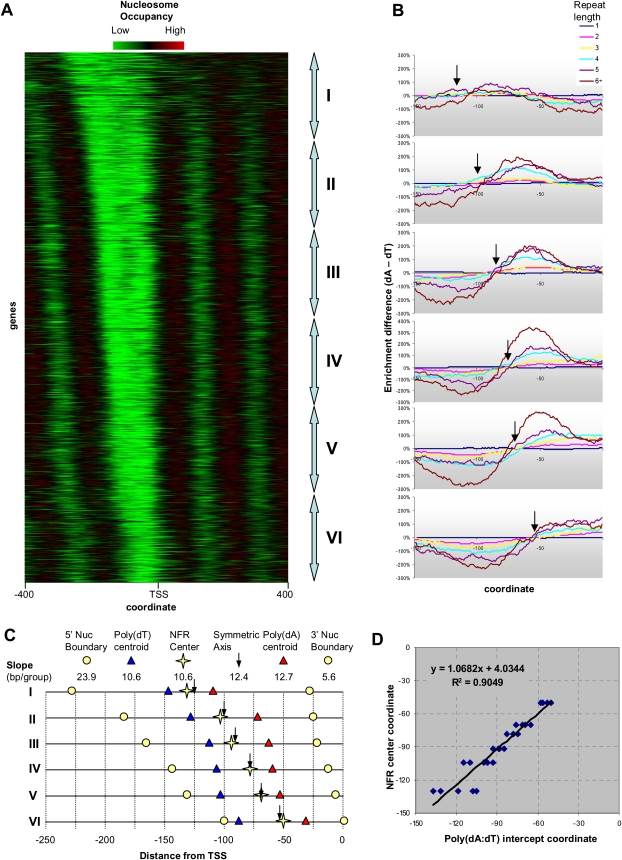
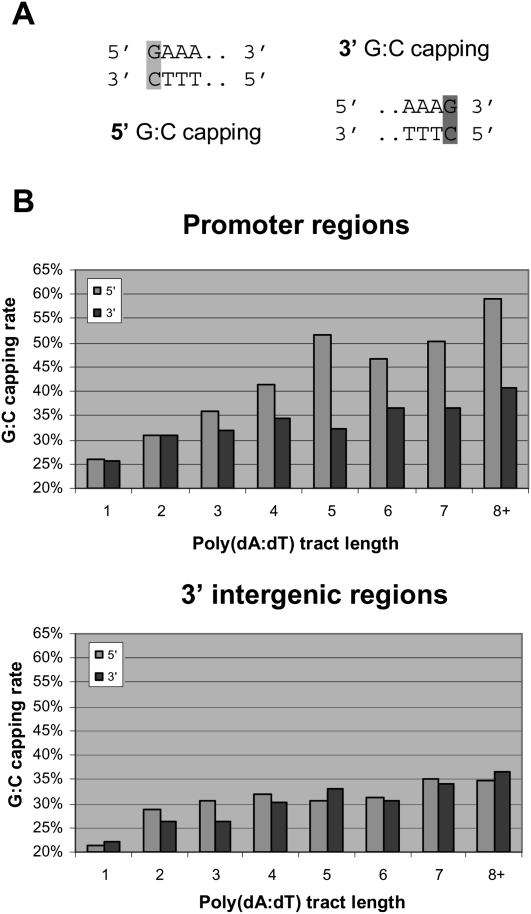
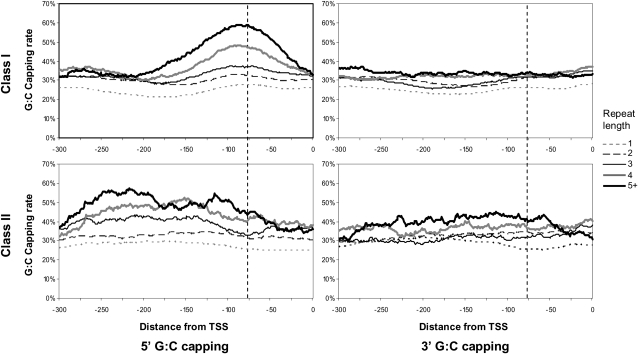
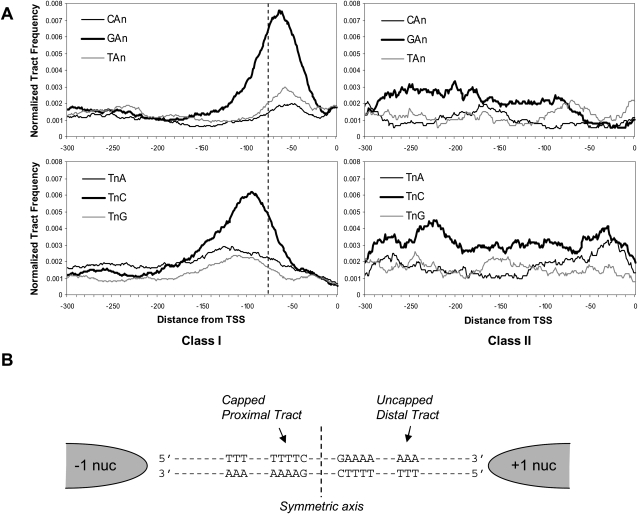
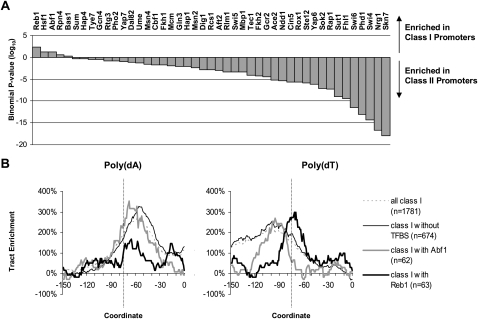
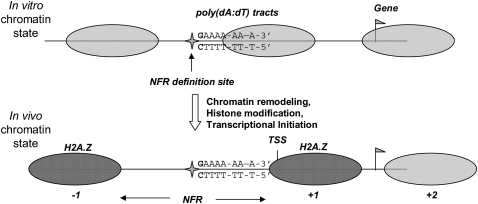
Eukaryotic transcriptional regulation is mediated by the organization of nucleosomes in promoter regions. Most Saccharomyces cerevisiae promoters have a highly stereotyped chromatin organization, where nucleosome-free regions (NFR) are flanked by well-ordered nucleosomes. We have found that yeast promoters fall into two classes differing in NFR sharpness, and that this distinction follows a known transcriptional dichotomy in yeast genes. A class of yeast promoters having well-defined NFRs are characterized by positioned patterns of poly(dA:dT) tracts with several novel features. First, poly(dA:dT) tracts are localized in a strand-dependent manner, with poly(dA) tracts lying proximal to transcriptional start sites and poly(dT) tracts lying distal, and collectively define a symmetry axis that is coincident with NFR centers. Second, poly(dA:dT) tracts are preferentially "capped" by G:C residues on the terminus proximal to the symmetry axis. Both signature features co-vary with fine positional variations between NFRs, establishing a closely knit relationship between poly(dA:dT) tracts, their capping patterns, and the central coordinates of NFRs. We found that these features are unique to promoters with well-defined NFRs, and that these promoters display significant difference between in vitro and in vivo nucleosome occupancy patterns. These observations are consistent with a model in which localized and G:C-capped poly(dA:dT) tracts initiate or facilitate the formation of NFRs at their center, possibly with chromatin remodeling and transcriptional machines involved.
Figures
References
-
- Albert I, Mavrich TN, Tomsho LP, Qi J, Zanton SJ, Schuster SC, Pugh BF. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature. 2007;446:572–576. - PubMed
-
- Bao Y, White CL, Luger K. Nucleosome core particles containing a poly(dA·dT) sequence element exhibit a locally distorted DNA structure. J Mol Biol. 2006;361:617–624. - PubMed
-
- Basehoar AD, Zanton SJ, Pugh BF. Identification and distinct regulation of yeast TATA box-containing genes. Cell. 2004;116:699–709. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases