

Cryo-EM model of the bullet-shaped vesicular stomatitis virus
- PMID: 20133572
- PMCID: PMC2892700
- DOI: 10.1126/science.1181766
Cryo-EM model of the bullet-shaped vesicular stomatitis virus
Erratum in
- Science. 2013 Feb 15;339(6121):757
Abstract
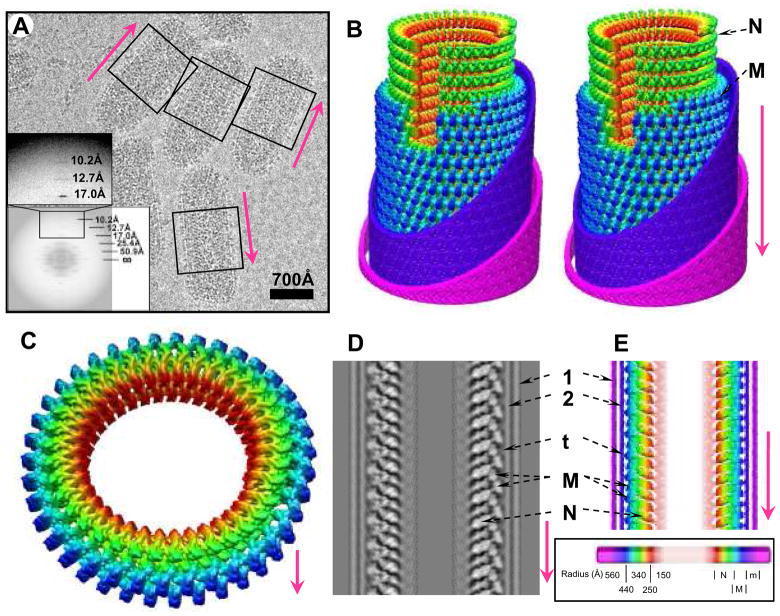
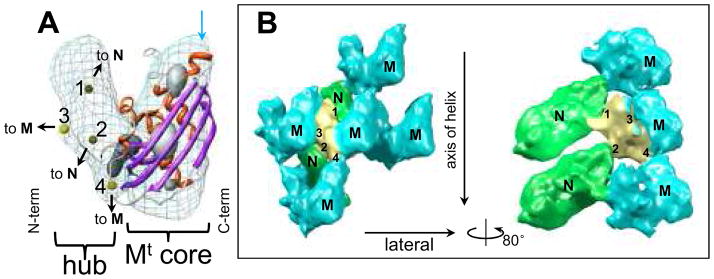
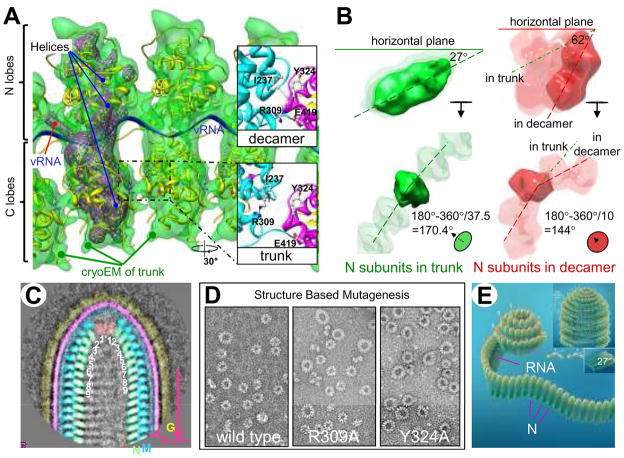
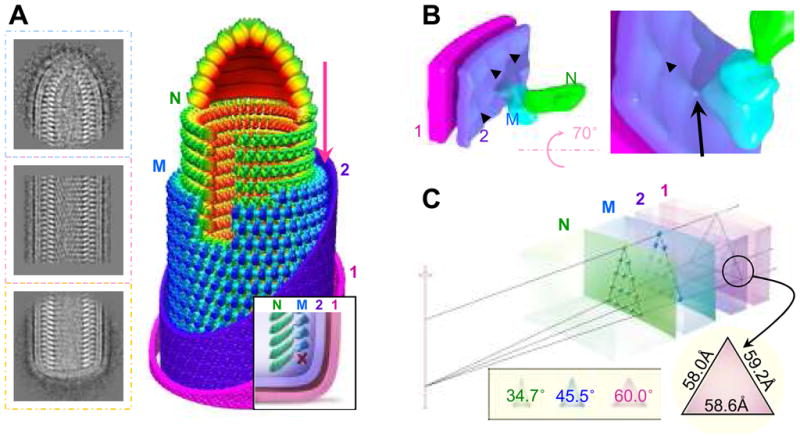
Vesicular stomatitis virus (VSV) is a bullet-shaped rhabdovirus and a model system of negative-strand RNA viruses. Through direct visualization by means of cryo-electron microscopy, we show that each virion contains two nested, left-handed helices: an outer helix of matrix protein M and an inner helix of nucleoprotein N and RNA. M has a hub domain with four contact sites that link to neighboring M and N subunits, providing rigidity by clamping adjacent turns of the nucleocapsid. Side-by-side interactions between neighboring N subunits are critical for the nucleocapsid to form a bullet shape, and structure-based mutagenesis results support this description. Together, our data suggest a mechanism of VSV assembly in which the nucleocapsid spirals from the tip to become the helical trunk, both subsequently framed and rigidified by the M layer.
Figures
References
-
- Lichty BD, Power AT, Stojdl DF, Bell JC. Trends Mol Med. 2004 May;10:210. - PubMed
-
- Stojdl DF, et al. Cancer Cell. 2003 Oct;4:263. - PubMed
-
- Schnell MJ, Johnson JE, Buonocore L, Rose JK. Cell. 1997 Sep 5;90:849. - PubMed
-
- Mebatsion T, Finke S, Weiland F, Conzelmann KK. Cell. 1997 Sep 5;90:841. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
