

Premature replacement of mu with alpha immunoglobulin chains impairs lymphopoiesis and mucosal homing but promotes plasma cell maturation
- PMID: 20133609
- PMCID: PMC2840347
- DOI: 10.1073/pnas.0912393107
Premature replacement of mu with alpha immunoglobulin chains impairs lymphopoiesis and mucosal homing but promotes plasma cell maturation
Abstract
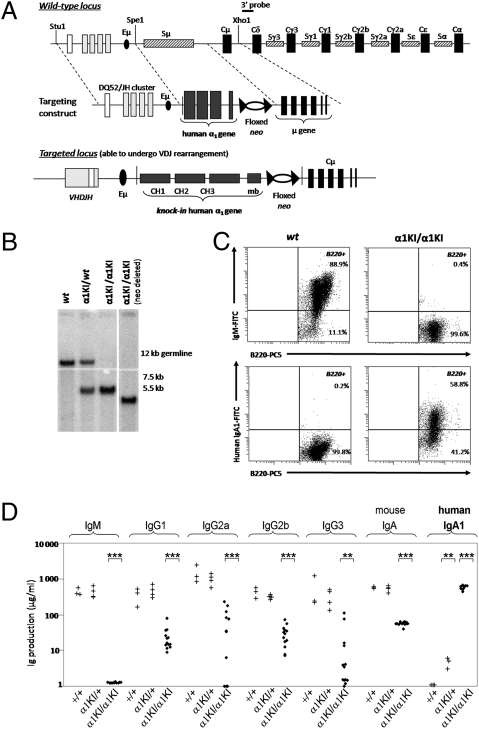
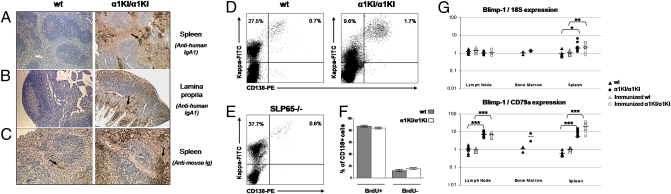
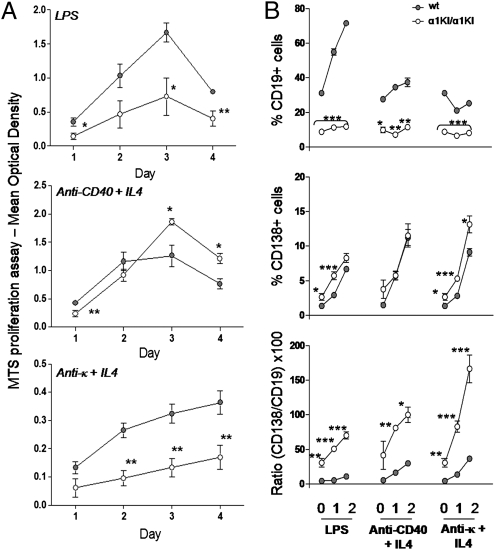
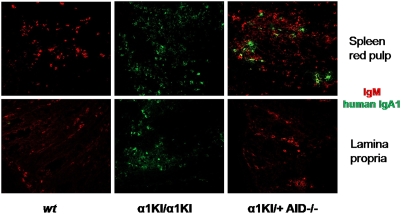
Sequentially along B cell differentiation, the different classes of membrane Ig heavy chains associate with the Ig alpha/Ig beta heterodimer within the B cell receptor (BCR). Whether each Ig class conveys specific signals adapted to the corresponding differentiation stage remains debated. We investigated the impact of the forced expression of an IgA-class receptor throughout murine B cell differentiation by knocking in the human C alpha Ig gene in place of the S mu region. Despite expression of a functional BCR, homozygous mutant mice showed a partial developmental blockade at the pro-B/pre-BI and large pre-BII cell stages, with decreased numbers of small pre-BII cells. Beyond this stage, peripheral B cell compartments of reduced size developed and allowed specific antibody responses, whereas mature cells showed constitutive activation and a strong commitment to plasma cell differentiation. Secreted IgA correctly assembled into polymers, associated with the murine J chain, and was transported into secretions. In heterozygous mutants, cells expressing the IgA allele competed poorly with those expressing IgM from the wild-type allele and were almost undetectable among peripheral B lymphocytes, notably in gut-associated lymphoid tissues. Our data indicate that the IgM BCR is more efficient in driving early B cell education and in mucosal site targeting, whereas the IgA BCR appears particularly suited to promoting activation and differentiation of effector plasma cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Brandtzaeg P. Induction of secretory immunity and memory at mucosal surfaces. Vaccine. 2007;25:5467–5484. - PubMed
-
- Flanagan JG, Lefranc MP, Rabbitts TH. Mechanisms of divergence and convergence of the human immunoglobulin α 1 and α 2 constant region gene sequences. Cell. 1984;36:681–688. - PubMed
-
- Cogné M, Preud’homme JL. Gene deletions force nonsecretory α-chain disease plasma cells to produce membrane-form α-chain only. J Immunol. 1990;145:2455–2458. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
