

An efficient, multiply promiscuous hydrolase in the alkaline phosphatase superfamily
- PMID: 20133613
- PMCID: PMC2840280
- DOI: 10.1073/pnas.0903951107
An efficient, multiply promiscuous hydrolase in the alkaline phosphatase superfamily
Abstract
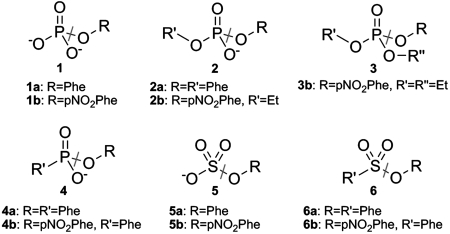
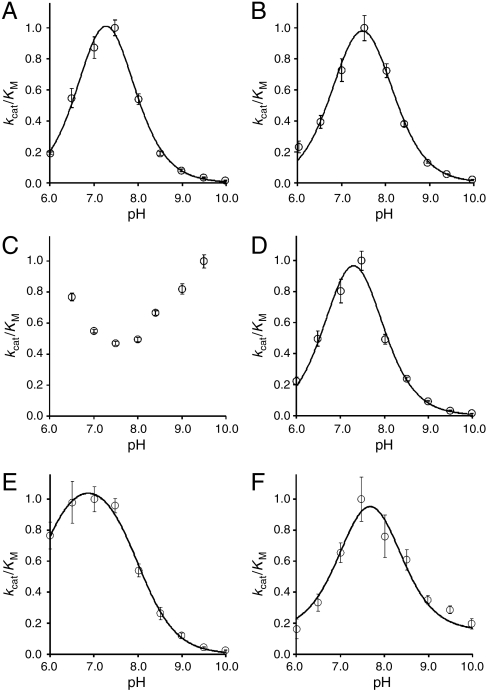
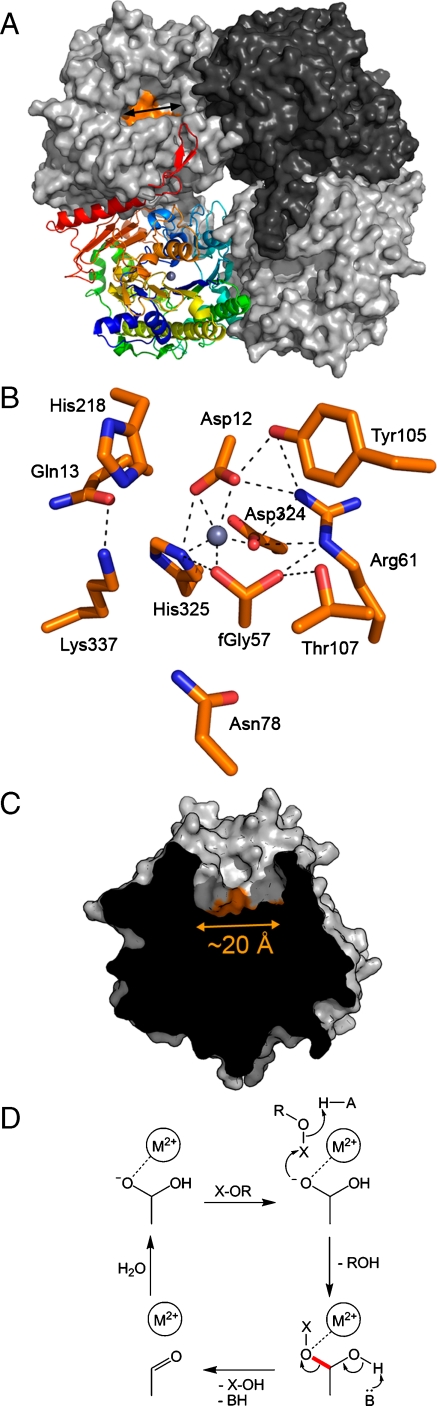
We report a catalytically promiscuous enzyme able to efficiently promote the hydrolysis of six different substrate classes. Originally assigned as a phosphonate monoester hydrolase (PMH) this enzyme exhibits substantial second-order rate accelerations ((k(cat)/K(M))/k(w)), ranging from 10(7) to as high as 10(19), for the hydrolyses of phosphate mono-, di-, and triesters, phosphonate monoesters, sulfate monoesters, and sulfonate monoesters. This substrate collection encompasses a range of substrate charges between 0 and -2, transition states of a different nature, and involves attack at two different reaction centers (P and S). Intrinsic reactivities (half-lives) range from 200 days to 10(5) years under near neutrality. The substantial rate accelerations for a set of relatively difficult reactions suggest that efficient catalysis is not necessarily limited to efficient stabilization of just one transition state. The crystal structure of PMH identifies it as a member of the alkaline phosphatase superfamily. PMH encompasses four of the native activities previously observed in this superfamily and extends its repertoire by two further activities, one of which, sulfonate monoesterase, has not been observed previously for a natural enzyme. PMH is thus one of the most promiscuous hydrolases described to date. The functional links between superfamily activities can be presumed to have played a role in functional evolution by gene duplication.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- O’Brien PJ, Herschlag D. Catalytic promiscuity and the evolution of new enzymatic activities. Chem Biol. 1999;6:R91–R105. - PubMed
-
- Khersonsky O, Roodveldt C, Tawfik DS. Enzyme promiscuity: evolutionary and mechanistic aspects. Curr Opin Chem Biol. 2006;10:498–508. - PubMed
-
- Jonas S, Hollfelder F. Mechanism and Catalytic Promiscuity: Emerging Mechanistic Principles for Identification and Manipulation of Catalytically Promiscuous Enzymes. In: Bornscheuer U, Lutz S, editors. Handbook of Protein Engineering. Vol 1. Weinheim, Germany: Wiley VCH; 2008. pp. 47–79.
-
- Glasner ME, Gerlt JA, Babbitt PC. Evolution of enzyme superfamilies. Curr Opin Chem Biol. 2006;10:492–497. - PubMed
-
- Aharoni A, et al. The ‘evolvability’ of promiscuous protein functions. Nat Genet. 2005;37:73–76. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
