

Structural insight into M-band assembly and mechanics from the titin-obscurin-like-1 complex
- PMID: 20133654
- PMCID: PMC2814874
- DOI: 10.1073/pnas.0913736107
Structural insight into M-band assembly and mechanics from the titin-obscurin-like-1 complex
Abstract
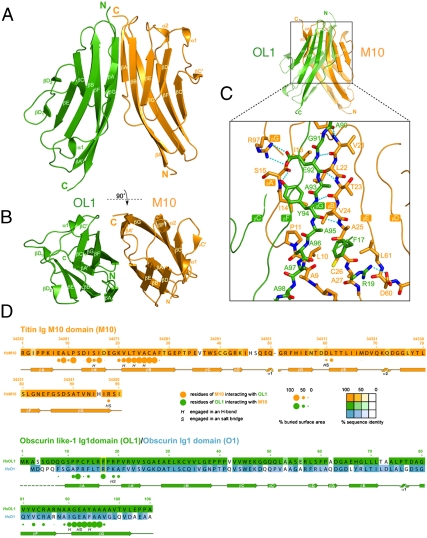
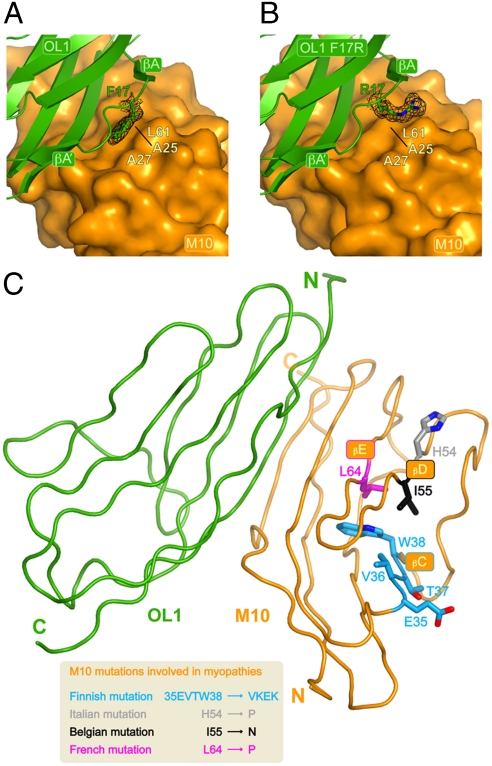
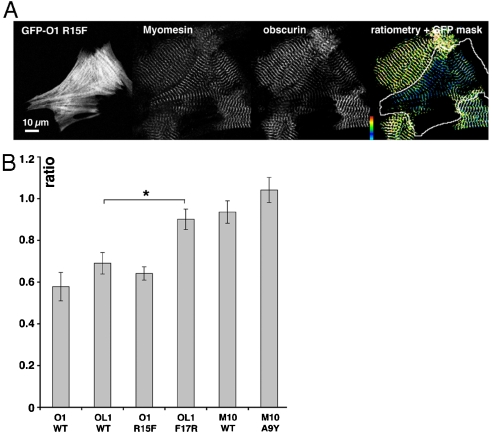
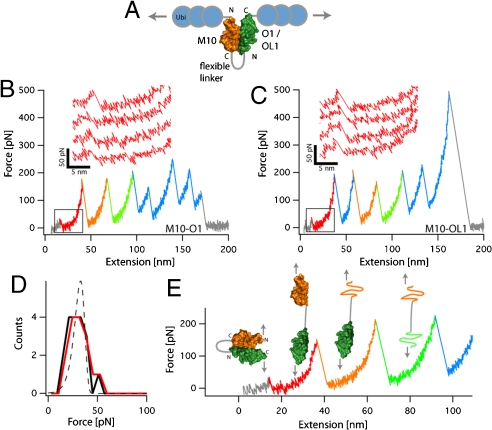
In the sarcomeric M-band, the giant ruler proteins titin and obscurin, its small homologue obscurin-like-1 (obsl1), and the myosin cross-linking protein myomesin form a ternary complex that is crucial for the function of the M-band as a mechanical link. Mutations in the last titin immunoglobulin (Ig) domain M10, which interacts with the N-terminal Ig-domains of obscurin and obsl1, lead to hereditary muscle diseases. The M10 domain is unusual not only in that it is a frequent target of disease-linked mutations, but also in that it is the only currently known muscle Ig-domain that interacts with two ligands--obscurin and obsl1--in different sarcomeric subregions. Using x-ray crystallography, we show the structural basis for titin M10 interaction with obsl1 in a novel antiparallel Ig-Ig architecture and unravel the molecular basis of titin-M10 linked myopathies. The severity of these pathologies correlates with the disruption of the titin-obsl1/obscurin complex. Conserved signature residues at the interface account for differences in affinity that direct the cellular sorting in cardiomyocytes. By engineering the interface signature residues of obsl1 to obscurin, and vice versa, their affinity for titin can be modulated similar to the native proteins. In single-molecule force-spectroscopy experiments, both complexes yield at forces of around 30 pN, much lower than those observed for the mechanically stable Z-disk complex of titin and telethonin, suggesting why even moderate weakening of the obsl1/obscurin-titin links has severe consequences for normal muscle functions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The crystal structure of the human titin:obscurin complex reveals a conserved yet specific muscle M-band zipper module.J Mol Biol. 2015 Feb 27;427(4):718-736. doi: 10.1016/j.jmb.2014.11.019. Epub 2014 Dec 6. J Mol Biol. 2015. PMID: 25490259
-
Interactions with titin and myomesin target obscurin and obscurin-like 1 to the M-band: implications for hereditary myopathies.J Cell Sci. 2008 Jun 1;121(11):1841-51. doi: 10.1242/jcs.028019. Epub 2008 May 13. J Cell Sci. 2008. PMID: 18477606
-
Binding of Myomesin to Obscurin-Like-1 at the Muscle M-Band Provides a Strategy for Isoform-Specific Mechanical Protection.Structure. 2017 Jan 3;25(1):107-120. doi: 10.1016/j.str.2016.11.015. Epub 2016 Dec 15. Structure. 2017. PMID: 27989621 Free PMC article.
-
The role of the M-band myomesin proteins in muscle integrity and cardiac disease.J Biomed Sci. 2022 Mar 7;29(1):18. doi: 10.1186/s12929-022-00801-6. J Biomed Sci. 2022. PMID: 35255917 Free PMC article. Review.
-
Fishing out proteins that bind to titin.J Cell Biol. 2001 Jul 9;154(1):21-4. doi: 10.1083/jcb.200106072. J Cell Biol. 2001. PMID: 11448986 Free PMC article. Review.
Cited by
-
Structure of giant muscle proteins.Front Physiol. 2013 Dec 12;4:368. doi: 10.3389/fphys.2013.00368. Front Physiol. 2013. PMID: 24376425 Free PMC article. Review.
-
Making sense of missense variants in TTN-related congenital myopathies.Acta Neuropathol. 2021 Mar;141(3):431-453. doi: 10.1007/s00401-020-02257-0. Epub 2021 Jan 15. Acta Neuropathol. 2021. PMID: 33449170 Free PMC article.
-
The role of single protein elasticity in mechanobiology.Nat Rev Mater. 2023 Jan;8:10-24. doi: 10.1038/s41578-022-00488-z. Epub 2022 Oct 24. Nat Rev Mater. 2023. PMID: 37469679 Free PMC article.
-
Three-dimensional structure of the M-region (bare zone) of vertebrate striated muscle myosin filaments by single-particle analysis.J Mol Biol. 2010 Nov 12;403(5):763-76. doi: 10.1016/j.jmb.2010.09.025. Epub 2010 Sep 17. J Mol Biol. 2010. PMID: 20851129 Free PMC article.
-
Increasing Role of Titin Mutations in Neuromuscular Disorders.J Neuromuscul Dis. 2016 Aug 30;3(3):293-308. doi: 10.3233/JND-160158. J Neuromuscul Dis. 2016. PMID: 27854229 Free PMC article. Review.
References
-
- Tskhovrebova L, Trinick J. Titin: Properties and family relationships. Nat Rev Mol Cell Biol. 2003;4:679–689. - PubMed
-
- Ehler E, Gautel M. Sarcomere and sarcomerogenesis. Exp Med Biol. 2008;642:1–14. - PubMed
-
- Lange S, Ehler E, Gautel M. From A to Z and back? Multicompartment proteins in the sarcomere. Trends Cell Biol. 2006;16:11–18. - PubMed
-
- Agarkova I, Perriard JC. The M-band: An elastic web that crosslinks thick filaments in the center of the sarcomere. Trends Cell Biol. 2005;15:477–485. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
