

The transition state for folding of an outer membrane protein
- PMID: 20133664
- PMCID: PMC2814873
- DOI: 10.1073/pnas.0911904107
The transition state for folding of an outer membrane protein
Abstract
Inspired by the seminal work of Anfinsen, investigations of the folding of small water-soluble proteins have culminated in detailed insights into how these molecules attain and stabilize their native folds. In contrast, despite their overwhelming importance in biology, progress in understanding the folding and stability of membrane proteins remains relatively limited. Here we use mutational analysis to describe the transition state involved in the reversible folding of the beta-barrel membrane protein PhoPQ-activated gene P (PagP) from a highly disordered state in 10 M urea to a native protein embedded in a lipid bilayer. Analysis of the equilibrium stability and unfolding kinetics of 19 variants that span all eight beta-strands of this 163-residue protein revealed that the transition-state structure is a highly polarized, partly formed beta-barrel. The results provide unique and detailed insights into the transition-state structure for beta-barrel membrane protein folding into a lipid bilayer and are consistent with a model for outer membrane protein folding via a tilted insertion mechanism.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
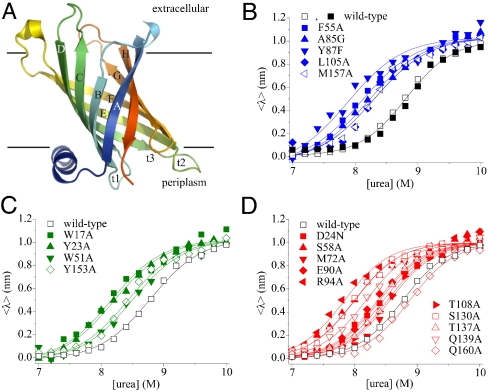
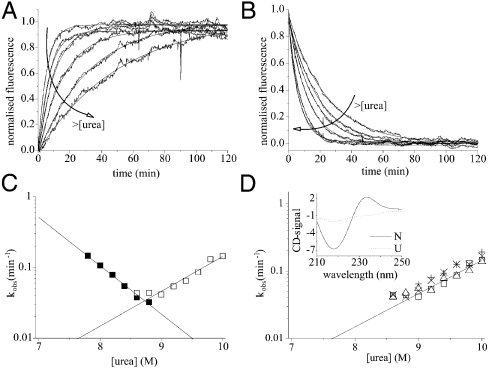
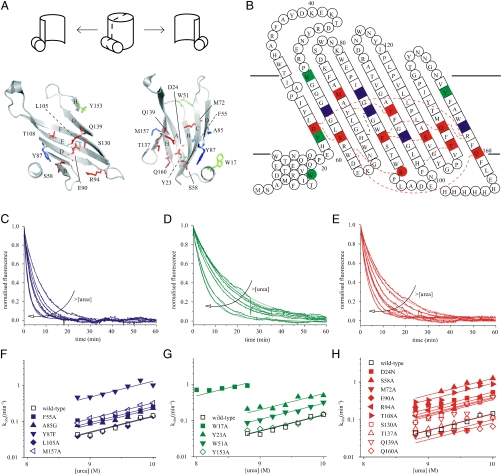
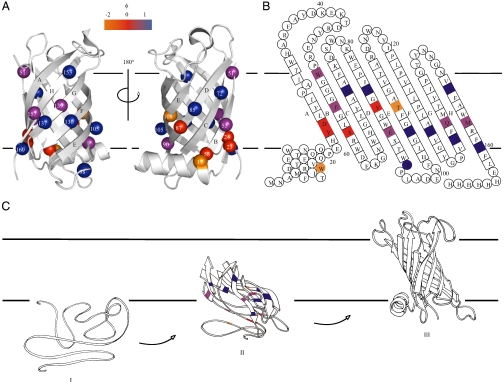
 [the cut-off generally accepted to yield highly reliable ΦF-values (41)].
[the cut-off generally accepted to yield highly reliable ΦF-values (41)].Comment in
-
Membrane protein folding makes the transition.Proc Natl Acad Sci U S A. 2010 Mar 2;107(9):3947-8. doi: 10.1073/pnas.0914478107. Epub 2010 Feb 19. Proc Natl Acad Sci U S A. 2010. PMID: 20173094 Free PMC article. No abstract available.
References
-
- Hediger MA, et al. The ABCs of solute carriers: Physiological, pathological and therapeutic implications of human membrane transport proteins. Pflugers Arch. 2004;447:465–468. - PubMed
-
- Sanders C, Myers JK. Disease-related misassembly of membrane proteins. Annu Rev Bioph Biom. 2004;33:25–51. - PubMed
-
- Chen GQ, Gouaux E. Probing the folding and unfolding of wild-type and mutant forms of bacteriorhodopsin in micellar solutions: Evaluation of reversible unfolding conditions. Biochemistry. 1999;38:15380–15387. - PubMed
-
- Bowie JU. Solving the membrane protein folding problem. Nature. 2005;438:581–589. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
