

Mechanical amplification by hair cells in the semicircular canals
- PMID: 20133682
- PMCID: PMC2840494
- DOI: 10.1073/pnas.0906765107
Mechanical amplification by hair cells in the semicircular canals
Abstract
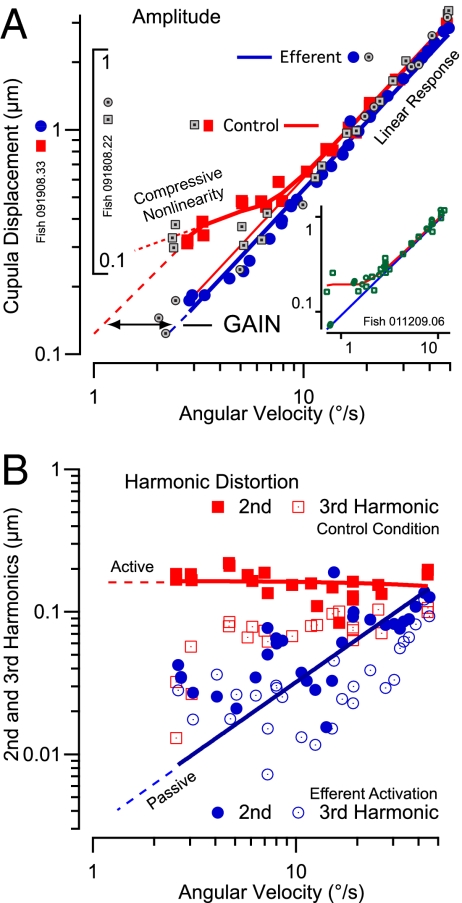
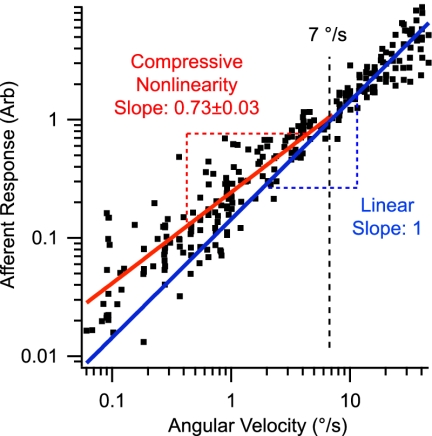
Sensory hair cells are the essential mechanotransducers of the inner ear, responsible not only for the transduction of sound and motion stimuli but also, remarkably, for nanomechanical amplification of sensory stimuli. Here we show that semicircular canal hair cells generate a mechanical nonlinearity in vivo that increases sensitivity to angular motion by amplification at low stimulus strengths. Sensitivity at high stimulus strengths is linear and shows no evidence of amplification. Results suggest that the mechanical work done by hair cells contributes approximately 97 zJ/cell of amplification per stimulus cycle, improving sensitivity to angular velocity stimuli below approximately 5 degrees /s (0.3-Hz sinusoidal motion). We further show that mechanical amplification can be inhibited by the brain via activation of efferent synaptic contacts on hair cells. The experimental model was the oyster toadfish, Opsanus tau. Physiological manifestation of mechanical amplification and efferent control in a teleost vestibular organ suggests the active motor process in sensory hair cells is ancestral. The biophysical basis of the motor(s) remains hypothetical, but a key discriminating question may involve how changes in somatic electrical impedance evoked by efferent synaptic action alter function of the motor(s).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
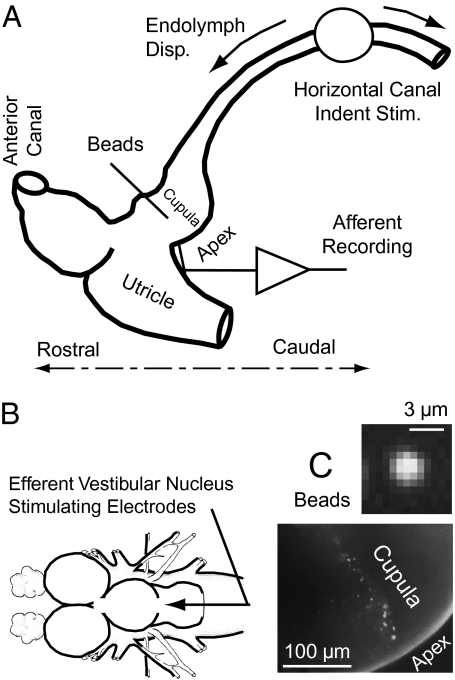
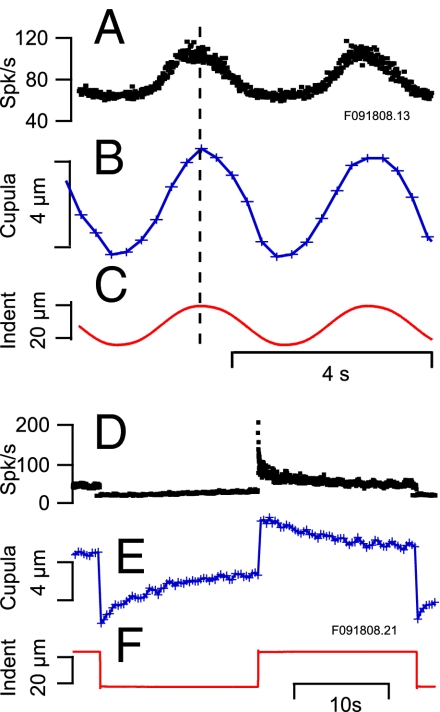
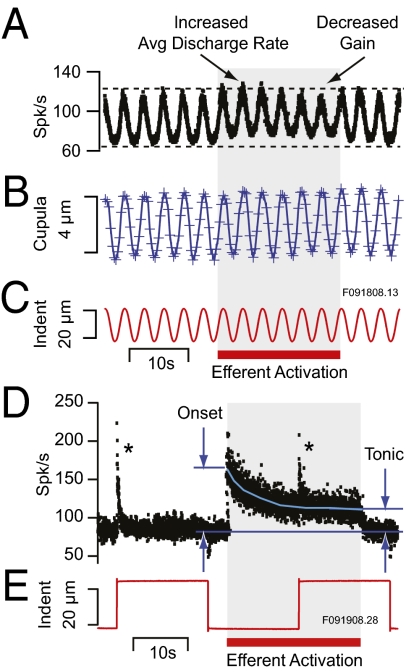
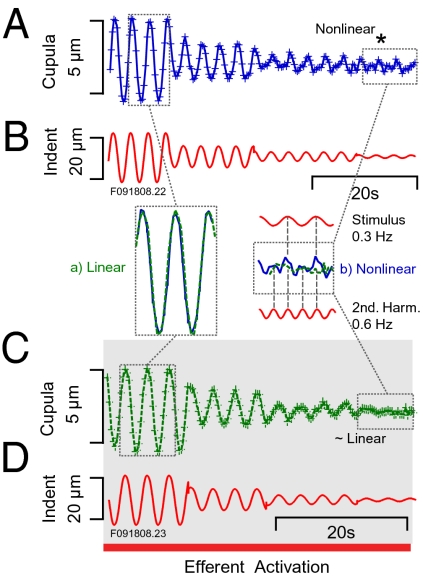
Similar articles
-
Efferent control of hair cell and afferent responses in the semicircular canals.J Neurophysiol. 2009 Sep;102(3):1513-25. doi: 10.1152/jn.91367.2008. Epub 2009 Jul 1. J Neurophysiol. 2009. PMID: 19571186 Free PMC article.
-
Relationship between inner-ear fluid pressure and semicircular canal afferent nerve discharge.J Assoc Res Otolaryngol. 2002 Mar;3(1):26-44. doi: 10.1007/s101620010088. J Assoc Res Otolaryngol. 2002. PMID: 12083722 Free PMC article.
-
Ultrastructural observations of efferent terminals in the crista Ampullaris of the toadfish, opsanus tau.Exp Brain Res. 2004 Apr;155(3):265-73. doi: 10.1007/s00221-003-1734-8. Epub 2003 Dec 19. Exp Brain Res. 2004. Corrected and republished in: Exp Brain Res. 2004 Jul;157(1):128-36. doi: 10.1007/s00221-004-1898-x. PMID: 14689144 Corrected and republished.
-
How endolymph pressure modulates semicircular canal primary afferent discharge.Ann N Y Acad Sci. 2001 Oct;942:313-21. doi: 10.1111/j.1749-6632.2001.tb03756.x. Ann N Y Acad Sci. 2001. PMID: 11710473 Review.
-
Molecular mechanisms and potentials for differentiating inner ear stem cells into sensory hair cells.Dev Biol. 2014 Jun 15;390(2):93-101. doi: 10.1016/j.ydbio.2014.03.010. Epub 2014 Mar 26. Dev Biol. 2014. PMID: 24680894 Review.
Cited by
-
Striated organelle, a cytoskeletal structure positioned to modulate hair-cell transduction.Proc Natl Acad Sci U S A. 2012 Mar 20;109(12):4473-8. doi: 10.1073/pnas.1101003109. Epub 2012 Mar 6. Proc Natl Acad Sci U S A. 2012. PMID: 22396594 Free PMC article.
-
The diverse effects of mechanical loading on active hair bundles.Proc Natl Acad Sci U S A. 2012 Feb 7;109(6):1943-8. doi: 10.1073/pnas.1120298109. Epub 2012 Jan 20. Proc Natl Acad Sci U S A. 2012. PMID: 22308449 Free PMC article.
-
Influence of Magnitude and Duration of Altered Gravity and Readaptation to 1 g on the Structure and Function of the Utricle in Toadfish, Opsanus tau.Front Physiol. 2018 Oct 22;9:1469. doi: 10.3389/fphys.2018.01469. eCollection 2018. Front Physiol. 2018. PMID: 30405430 Free PMC article.
-
Wave Mechanics of the Vestibular Semicircular Canals.Biophys J. 2017 Sep 5;113(5):1133-1149. doi: 10.1016/j.bpj.2017.08.001. Biophys J. 2017. PMID: 28877495 Free PMC article.
-
ACh-induced hyperpolarization and decreased resistance in mammalian type II vestibular hair cells.J Neurophysiol. 2018 Jan 1;119(1):312-325. doi: 10.1152/jn.00030.2017. Epub 2017 Oct 4. J Neurophysiol. 2018. PMID: 28978760 Free PMC article.
References
-
- Stensiö E. The Downtonian and Devonian vertebrates of Spitsbergen. Part I, Family Cephalaspidae. Skr . Svalbard Nordishavet. 1927;12:1–391.
-
- Brownell WE, Bader CRB, Bertrand D, de Ribaupierre Y. Evoked mechanical responses of isolated cochlear outer hair cells. Science. 1985;227:194–196. - PubMed
-
- Zheng J, et al. Prestin is the motor protein of cochlear outer hair cells. Nature. 2000;405:149–155. - PubMed
-
- Liberman MC, et al. Prestin is required for electromotility of the outer hair cell and for the cochlear amplifier. Nature. 2002;419:300–304. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
