

Targeted disruption of the CREB coactivator Crtc2 increases insulin sensitivity
- PMID: 20133702
- PMCID: PMC2840317
- DOI: 10.1073/pnas.0914897107
Targeted disruption of the CREB coactivator Crtc2 increases insulin sensitivity
Abstract
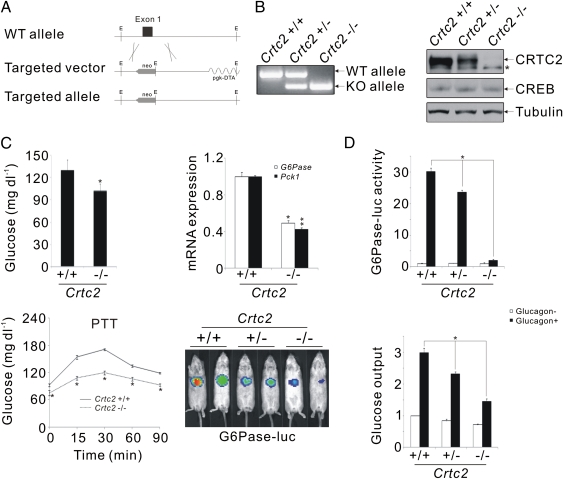
Under fasting conditions, increases in circulating concentrations of pancreatic glucagon maintain glucose homeostasis through induction of gluconeogenic genes by the CREB coactivator CRTC2. Hepatic CRTC2 activity is elevated in obesity, although the extent to which this cofactor contributes to attendant increases in insulin resistance is unclear. Here we show that mice with a knockout of the CRTC2 gene have decreased circulating glucose concentrations during fasting, due to attenuation of the gluconeogenic program. CRTC2 was found to stimulate hepatic gene expression in part through an N-terminal CREB binding domain that enhanced CREB occupancy over relevant promoters in response to glucagon. Deletion of sequences encoding the CREB binding domain in CRTC2 (-/-) mice lowered circulating blood glucose concentrations and improved insulin sensitivity in the context of diet-induced obesity. Our results suggest that small molecules that attenuate the CREB-CRTC2 pathway may provide therapeutic benefit to individuals with type 2 diabetes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Saltiel AR, Kahn CR. Insulin signalling and the regulation of glucose and lipid metabolism. Nature. 2001;414:799–806. - PubMed
-
- Saltiel AR. New perspectives into the molecular pathogenesis and treatment of type 2 diabetes. Cell. 2001;104:517–529. - PubMed
-
- Biddinger SB, Kahn CR. From mice to men: Insights into the insulin resistance syndromes. Annu Rev Physiol. 2006;68:123–158. - PubMed
-
- Koo SH, et al. The CREB coactivator TORC2 is a key regulator of fasting glucose metabolism. Nature. 2005;437:1109–1111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
