

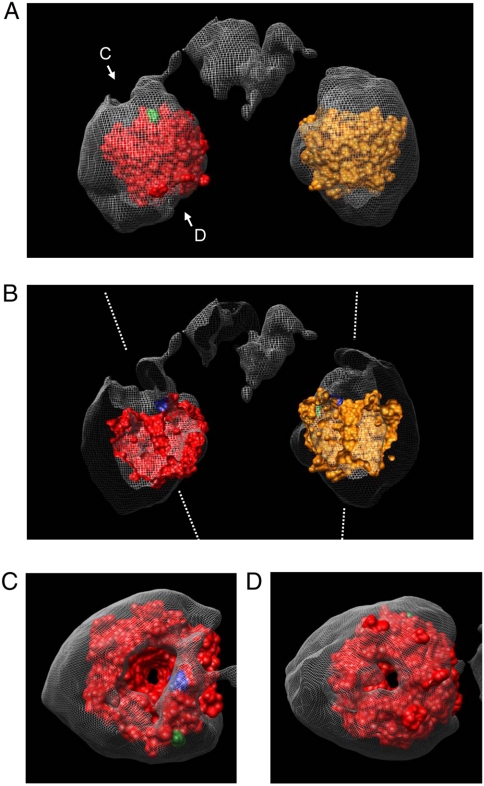
Keap1 is a forked-stem dimer structure with two large spheres enclosing the intervening, double glycine repeat, and C-terminal domains
- PMID: 20133743
- PMCID: PMC2840362
- DOI: 10.1073/pnas.0914036107
Keap1 is a forked-stem dimer structure with two large spheres enclosing the intervening, double glycine repeat, and C-terminal domains
Abstract
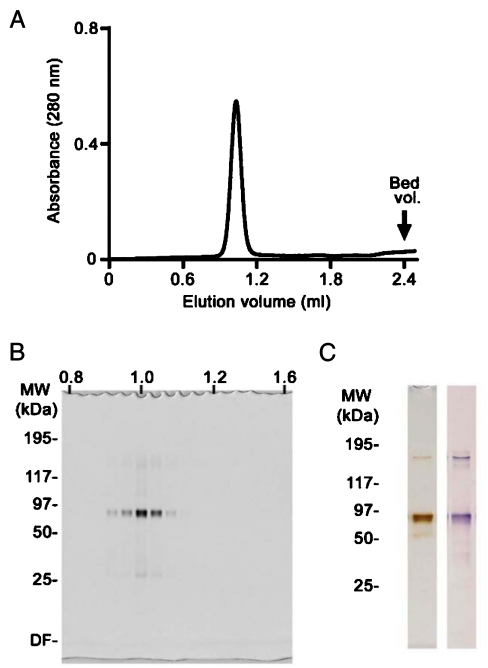
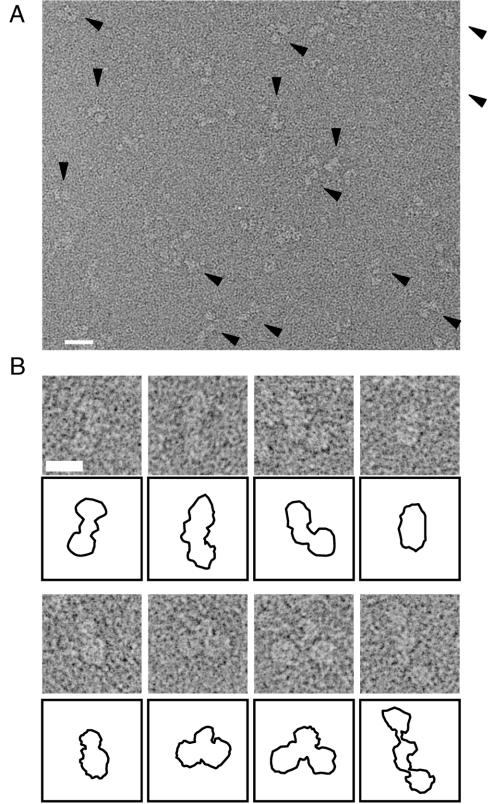
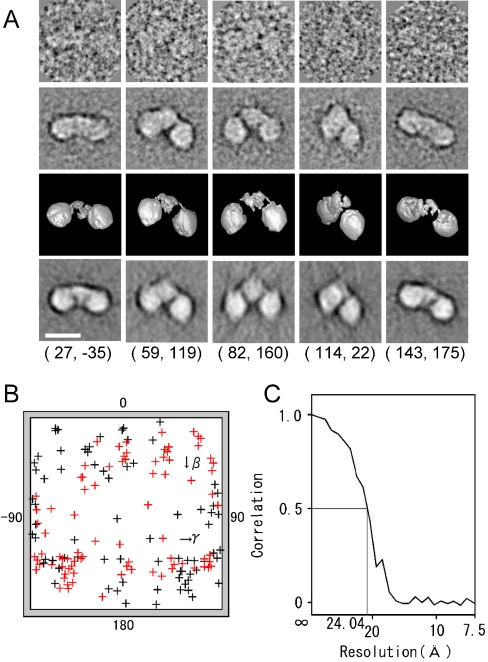
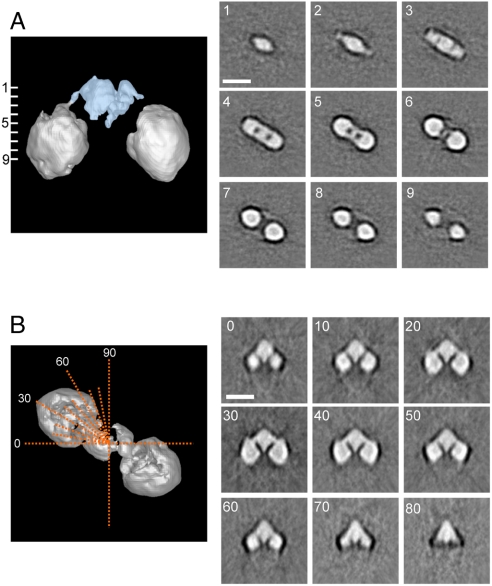
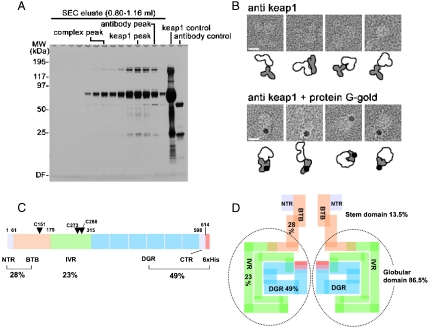
Keap1 is a substrate adaptor of a Cullin 3-based E3 ubiquitin ligase complex that recognizes Nrf2, and also acts as a cellular sensor for xenobiotics and oxidative stresses. Nrf2 is a transcriptional factor regulating the expression of cytoprotective enzyme genes in response to such stresses. Under unstressed conditions Keap1 binds Nrf2 and results in rapid degradation of Nrf2 through the proteasome pathway. In contrast, upon exposure to oxidative and electrophilic stress, reactive cysteine residues in intervening region (IVR) and Broad complex, Tramtrack, and Bric-à-Brac domains of Keap1 are modified by electrophiles. This modification prevents Nrf2 from rapid degradation and induces Nrf2 activity by repression of Keap1. Here we report the structure of mouse Keap1 homodimer by single particle electron microscopy. Three-dimensional reconstruction at 24-A resolution revealed two large spheres attached by short linker arms to the sides of a small forked-stem structure, resembling a cherry-bob. Each sphere has a tunnel corresponding to the central hole of the beta-propeller domain, as determined by x-ray crystallography. The IVR domain appears to surround the core of the beta-propeller domain. The unexpected proximity of IVR to the beta-propeller domain suggests that any distortions generated during modification of reactive cysteine residues in the IVR domain may send a derepression signal to the beta-propeller domain and thereby stabilize Nrf2. This study thus provides a structural basis for the two-site binding and hinge-latch model of stress sensing by the Nrf2-Keap1 system.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Itoh K, et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem Biophys Res Commun. 1997;236:313–322. - PubMed
-
- Kobayashi M, Yamamoto M. Nrf2-Keap1 regulation of cellular defense mechanisms against electrophiles and reactive oxygen species. Adv Enzyme Regul. 2006;46:113–140. - PubMed
-
- Wakabayashi N, et al. Keap1-null mutation leads to postnatal lethality due to constitutive Nrf2 activation. Nat Genet. 2003;35:238–245. - PubMed
-
- Okawa H, et al. Hepatocyte-specific deletion of the keap1 gene activates Nrf2 and confers potent resistance against acute drug toxicity. Biochem Biophys Res Commun. 2006;339:79–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
