

Synergistic antitumor effects of combined cathepsin B and cathepsin Z deficiencies on breast cancer progression and metastasis in mice
- PMID: 20133781
- PMCID: PMC2823914
- DOI: 10.1073/pnas.0907240107
Synergistic antitumor effects of combined cathepsin B and cathepsin Z deficiencies on breast cancer progression and metastasis in mice
Abstract
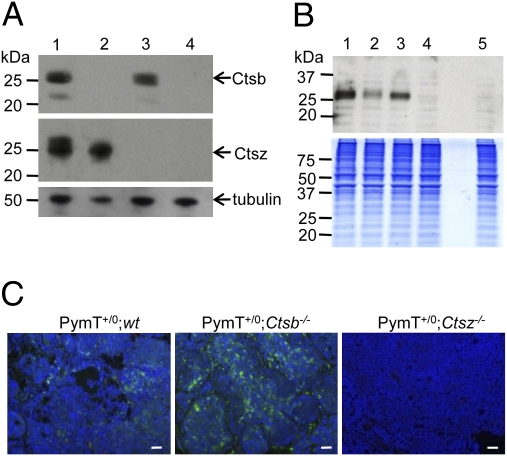
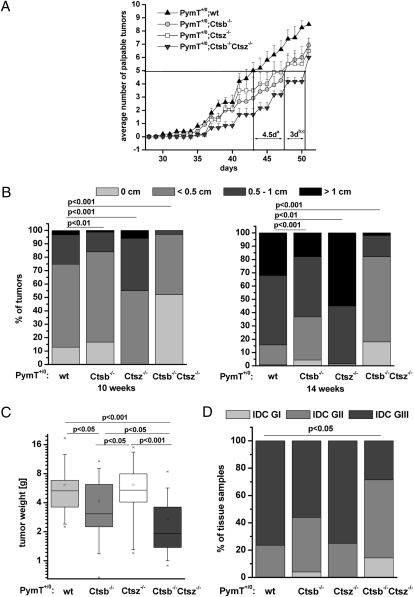
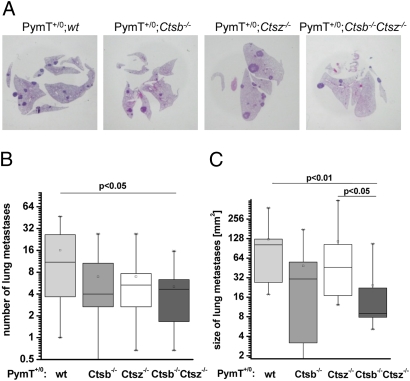
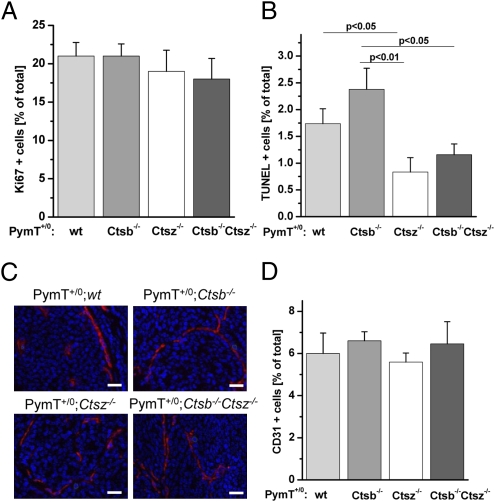
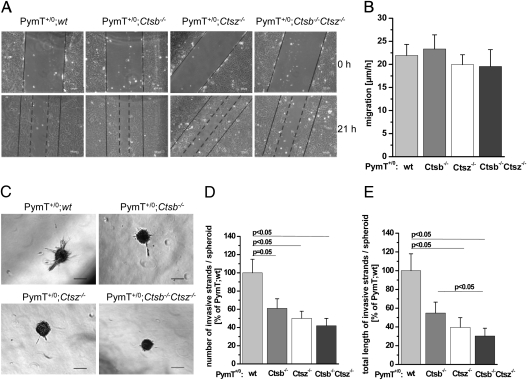
The lysosomal cysteine proteases cathepsin B (Ctsb) and cathepsin Z (Ctsz, also called cathepsin X/P) have been implicated in cancer pathogenesis. Compensation of Ctsb by Ctsz in Ctsb (-/-) mice has been suggested. To further define the functional interplay of these proteases in the context of cancer, we generated Ctsz null mice, crossed them with Ctsb-deficient mice harboring a transgene for the mammary duct-specific expression of polyoma middle T oncogene (PymT), and analyzed the effects of single and combined Ctsb and Ctsz deficiencies on breast cancer progression. Single Ctsb deficiency resulted in delayed detection of first tumors and reduced tumor burden, whereas Ctsz-deficient mice had only a prolonged tumor-free period. However, only a trend toward reduced metastatic burden without statistical significance was detected in both single mutants. Strikingly, combined loss of Ctsb and Ctsz led to additive effects, resulting in significant and prominent delay of early and advanced tumor development, improved histopathologic tumor grading, as well as a 70% reduction in the number of lung metastases and an 80% reduction in the size of these metastases. We conclude that the double deficiency of Ctsb and Ctsz exerts significant synergistic anticancer effects, whereas the single deficiencies demonstrate at least partial reciprocal compensation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Werb Z, Vu TH, Rinkenberger JL, Coussens LM. Matrix-degrading proteases and angiogenesis during development and tumor formation. APMIS. 1999;107:11–18. - PubMed
-
- Van Damme J, Struyf S, Opdenakker G. Chemokine–protease interactions in cancer. Semin Cancer Biol. 2004;14:201–208. - PubMed
-
- Bachmeier BE, Nerlich AG, Lichtinghagen R, Sommerhoff CP. Matrix metalloproteinases (MMPs) in breast cancer cell lines of different tumorigenicity. Anticancer Res. 2001;21(6A):3821–3828. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
