

T7 phage protein Gp2 inhibits the Escherichia coli RNA polymerase by antagonizing stable DNA strand separation near the transcription start site
- PMID: 20133868
- PMCID: PMC2836649
- DOI: 10.1073/pnas.0907908107
T7 phage protein Gp2 inhibits the Escherichia coli RNA polymerase by antagonizing stable DNA strand separation near the transcription start site
Abstract
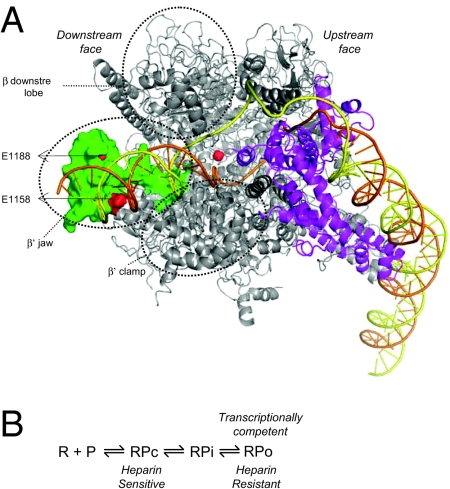
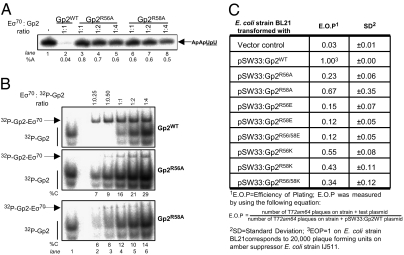
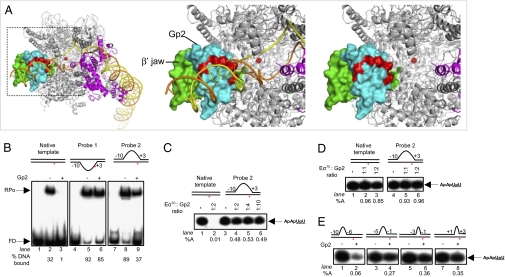
Infection of Escherichia coli by the T7 phage leads to rapid and selective inhibition of the host RNA polymerase (RNAP)--a multi-subunit enzyme responsible for gene transcription--by a small ( approximately 7 kDa) phage-encoded protein called Gp2. Gp2 is also a potent inhibitor of E. coli RNAP in vitro. Here we describe the first atomic resolution structure of Gp2, which reveals a distinct run of surface-exposed negatively charged amino acid residues on one side of the molecule. Our comprehensive mutagenesis data reveal that two conserved arginine residues located on the opposite side of Gp2 are important for binding to and inhibition of RNAP. Based on a structural model of the Gp2-RNAP complex, we propose that inhibition of transcription by Gp2 involves prevention of RNAP-promoter DNA interactions required for stable DNA strand separation and maintenance of the "transcription bubble" near the transcription start site, an obligatory step in the formation of a transcriptionally competent promoter complex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R01 GM059295/GM/NIGMS NIH HHS/United States
- GM59295/GM/NIGMS NIH HHS/United States
- BB/E023703/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E023703/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0501703/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
