

Phospholipase C activity affinity purifies with the Torpedo nicotinic acetylcholine receptor
- PMID: 20133947
- PMCID: PMC2856239
- DOI: 10.1074/jbc.M109.071993
Phospholipase C activity affinity purifies with the Torpedo nicotinic acetylcholine receptor
Abstract
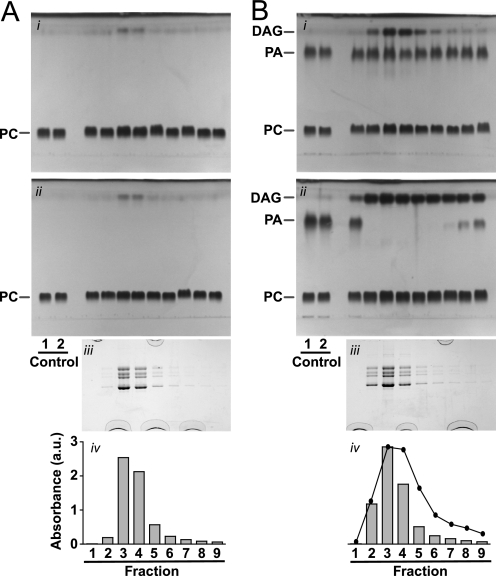
Nicotinic acetylcholine receptors mediate fast synaptic transmission by fluxing ions across the membrane in response to neurotransmitter binding. We show here that during affinity purification of the nicotinic acetylcholine receptor from Torpedo, phosphatidic acid, but not other anionic or zwitterionic phospholipids, is hydrolyzed to diacylglycerol. The phospholipase C activity elutes with the acetylcholine receptor and is inhibited by a lipid phosphate phosphohydrolase inhibitor, sodium vanadate, but not a phosphatidate phosphohydrolase inhibitor, N-ethylmaleimide. Further, the hydrolysis product of phosphatidic acid, diacylglycerol, enhances the functional capabilities of the acetylcholine receptor in the presence of anionic lipids. We conclude that a phospholipase C activity, which appears to be specific for phosphatidic acid, is associated with the nicotinic acetylcholine receptor. The acetylcholine receptor may directly or indirectly influence lipid metabolism in a manner that enhances its own function.
Figures




Similar articles
-
Correlation of phospholipid structure with functional effects on the nicotinic acetylcholine receptor. A modulatory role for phosphatidic acid.Biophys J. 1993 Mar;64(3):716-23. doi: 10.1016/S0006-3495(93)81431-0. Biophys J. 1993. PMID: 8471723 Free PMC article.
-
Phospholipase A2 hydrolysis of membrane phospholipids causes structural alteration of the nicotinic acetylcholine receptor.Biochim Biophys Acta. 1988 Feb 8;938(1):35-43. doi: 10.1016/0005-2736(88)90119-8. Biochim Biophys Acta. 1988. PMID: 3337815
-
Lipid composition alters drug action at the nicotinic acetylcholine receptor.Mol Pharmacol. 2008 Mar;73(3):880-90. doi: 10.1124/mol.107.039008. Epub 2007 Nov 30. Mol Pharmacol. 2008. PMID: 18055762
-
Immunisation with Torpedo acetylcholine receptor.Prog Neurobiol. 1984;23(1-2):39-62. doi: 10.1016/0301-0082(84)90011-x. Prog Neurobiol. 1984. PMID: 6097937 Review.
-
Nicotinic receptor of acetylcholine: structure of an oligomeric integral membrane protein.Physiol Rev. 1984 Oct;64(4):1162-239. doi: 10.1152/physrev.1984.64.4.1162. Physiol Rev. 1984. PMID: 6208568 Review. No abstract available.
Cited by
-
Tools for Understanding Nanoscale Lipid Regulation of Ion Channels.Trends Biochem Sci. 2019 Sep;44(9):795-806. doi: 10.1016/j.tibs.2019.04.001. Epub 2019 May 3. Trends Biochem Sci. 2019. PMID: 31060927 Free PMC article. Review.
-
A distinct mechanism for activating uncoupled nicotinic acetylcholine receptors.Nat Chem Biol. 2013 Nov;9(11):701-7. doi: 10.1038/nchembio.1338. Epub 2013 Sep 8. Nat Chem Biol. 2013. PMID: 24013278
-
Cations mediate interactions between the nicotinic acetylcholine receptor and anionic lipids.Biophys J. 2010 Mar 17;98(6):989-98. doi: 10.1016/j.bpj.2009.11.030. Biophys J. 2010. PMID: 20303856 Free PMC article.
-
Molecular mechanisms of acetylcholine receptor-lipid interactions: from model membranes to human biology.Biophys Rev. 2013 Mar;5(1):1-9. doi: 10.1007/s12551-012-0078-7. Epub 2012 May 10. Biophys Rev. 2013. PMID: 28510176 Free PMC article. Review.
-
A Closely Associated Phospholipase C Regulates Cation Channel Function through Phosphoinositide Hydrolysis.J Neurosci. 2018 Aug 29;38(35):7622-7634. doi: 10.1523/JNEUROSCI.0586-18.2018. Epub 2018 Jul 23. J Neurosci. 2018. PMID: 30037836 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
