

Review
doi: 10.1152/physiol.00039.2009.
Voltage-gated proton channels find their dream job managing the respiratory burst in phagocytes
Affiliations
- PMID: 20134026
- PMCID: PMC3023998
- DOI: 10.1152/physiol.00039.2009
Item in Clipboard
Review
Voltage-gated proton channels find their dream job managing the respiratory burst in phagocytes
Physiology (Bethesda).
2010 Feb.
Abstract
The voltage-gated proton channel bears surprising resemblance to the voltage-sensing domain (S1-S4) of other voltage-gated ion channels but is a dimer with two conduction pathways. The proton channel seems designed for efficient proton extrusion from cells. In phagocytes, it facilitates the production of reactive oxygen species by NADPH oxidase.
Figures
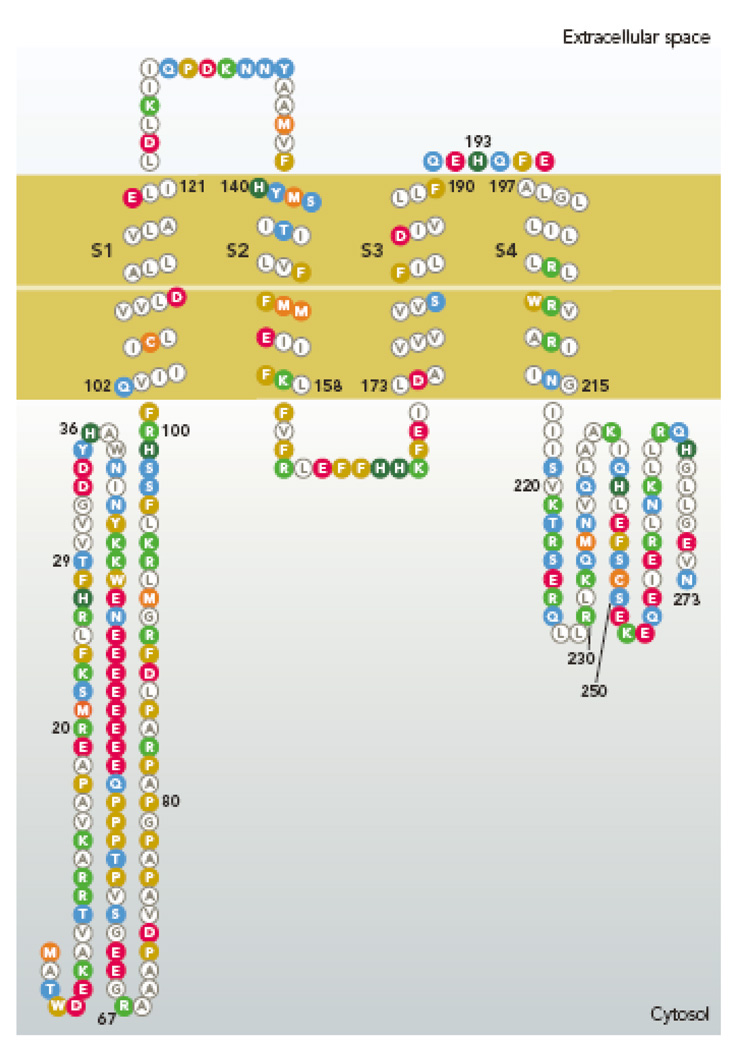
Amino acid sequence of the human proton channel, HV1 (Hvcn1 gene product). The four membrane spanning regions resemble S1–S4 of K+ channels, including 3 of the 4 Arg residues in S4 that are thought to sense voltage (173). Membrane segments were determined by refined PHD prediction (179). [Image created with TOPO2 software from the Sequence Analysis & Consulting Service at UCSF, then redrawn.]
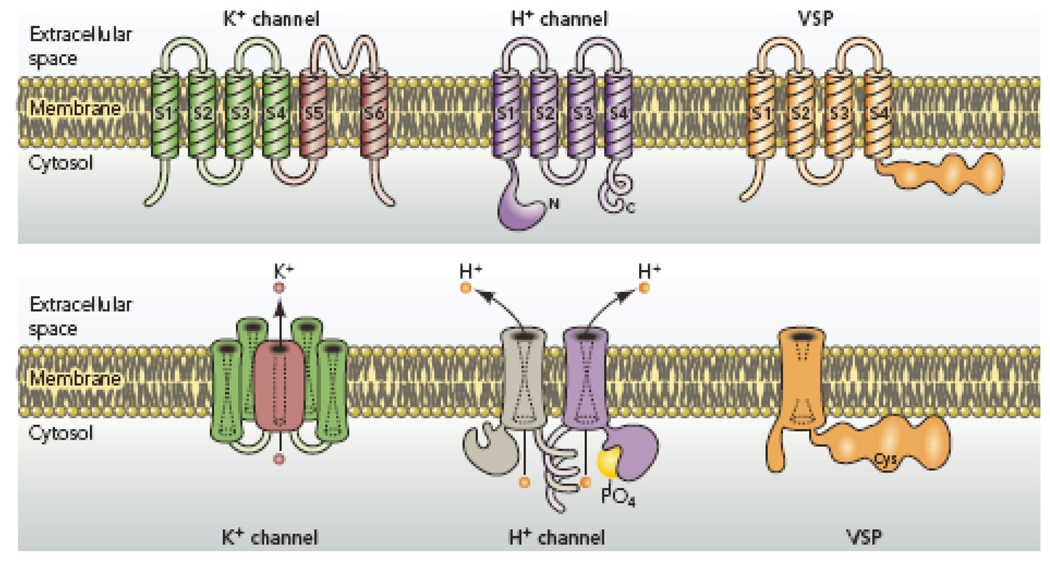
Comparison of architectural features of K+ channels, H+ channels, and voltage-sensing phosphatases (VSP). K+ channels are tetramers of subunits that each contain 6 membrane spanning regions, of which S1–S4 comprise the voltage sensing domain (VSD) and S5–S6 form the pore. The S5–S6 regions from each subunit together form a single central pore, which is surrounded by four VSDs. The voltage gated proton channel contains S1–S4 regions that are quite similar to the K+ channel VSD, but lacks the pore domain S5–S6 (173, 184). The proton channel is a dimer in which each monomer has its own conduction pathway (107, 116, 212). The main site of attachment is the C terminal coiled-coil domain (107, 116, 212). The N terminal intracellular region contains a phosphorylation site thought to convert the channel from “resting mode” to “enhanced gating mode” (56, 136, 152). The voltage sensing phosphatase shares similar S1–S4 regions with the others, but lacks conduction. Instead, it senses membrane potential and modulates phosphatase enzyme activity accordingly (147, 148).
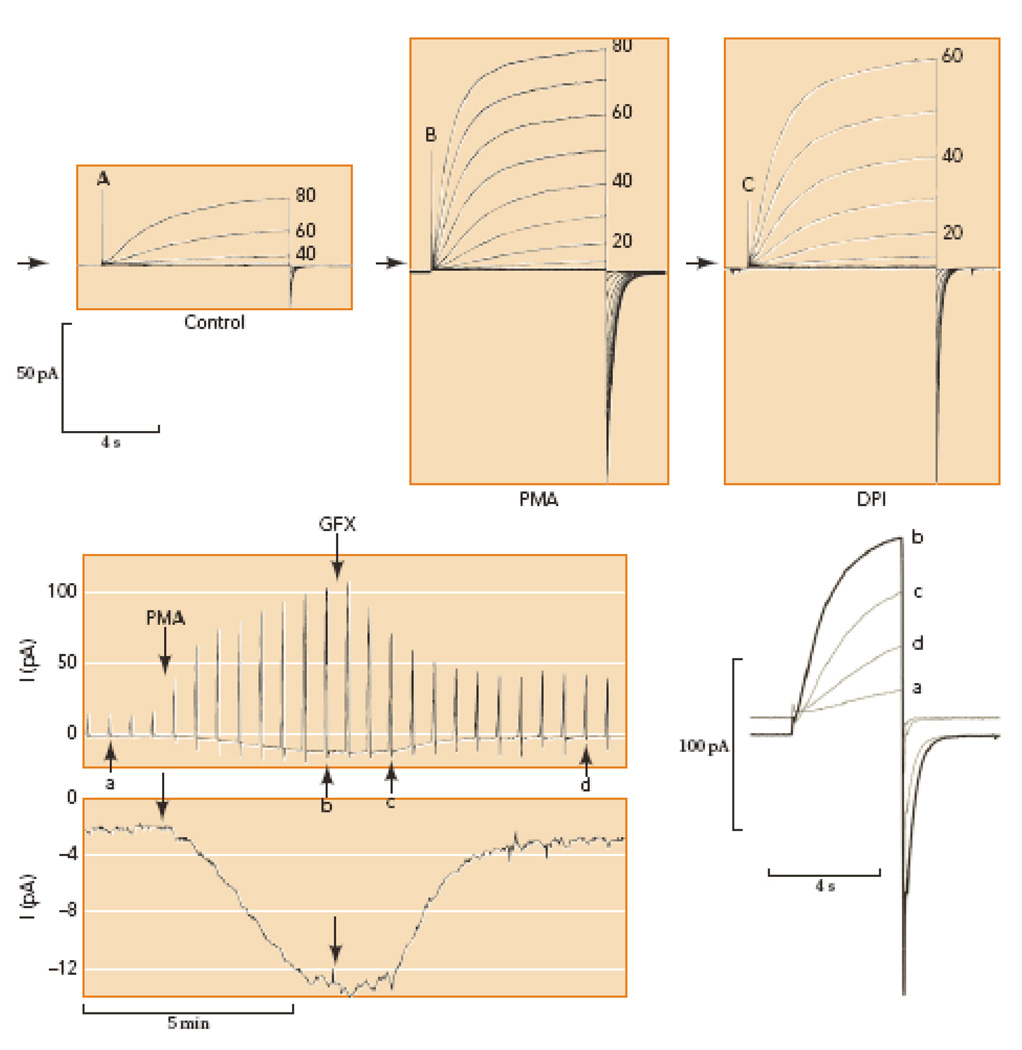
Conversion of proton channels in human eosinophils from resting mode (A) to enhanced gating mode (B) by PMA stimulation in perforated-patch configuration. Families of pulses were applied from a holding potential of −60 mV in 20-mV (A) or 10-mV steps (B–C) as indicated. Arrows indicate zero current; note the small inward electron current elicited by PMA (B). Inhibition of NADPH oxidase with diphenylene iodonium, DPI (C) eliminates the inward electron current, without immediate effect on the proton current. Lower panel shows the time course of the responses to PMA and GFX in a different eosinophil. Outward proton currents during pulses to +60 mV every 30 s are shown (above), with individual records identified by lower-case letters superimposed on the right at an expanded time scale. Net inward current at −60 mV at higher gain (below), reflects electron current through NADPH oxidase. PMA activates PKC and GFX inhibits PKC; thus, enhanced proton channel gating and NADPH oxidase are both regulated by phosphorylation. [Taken from 54 and 136, with permission.]
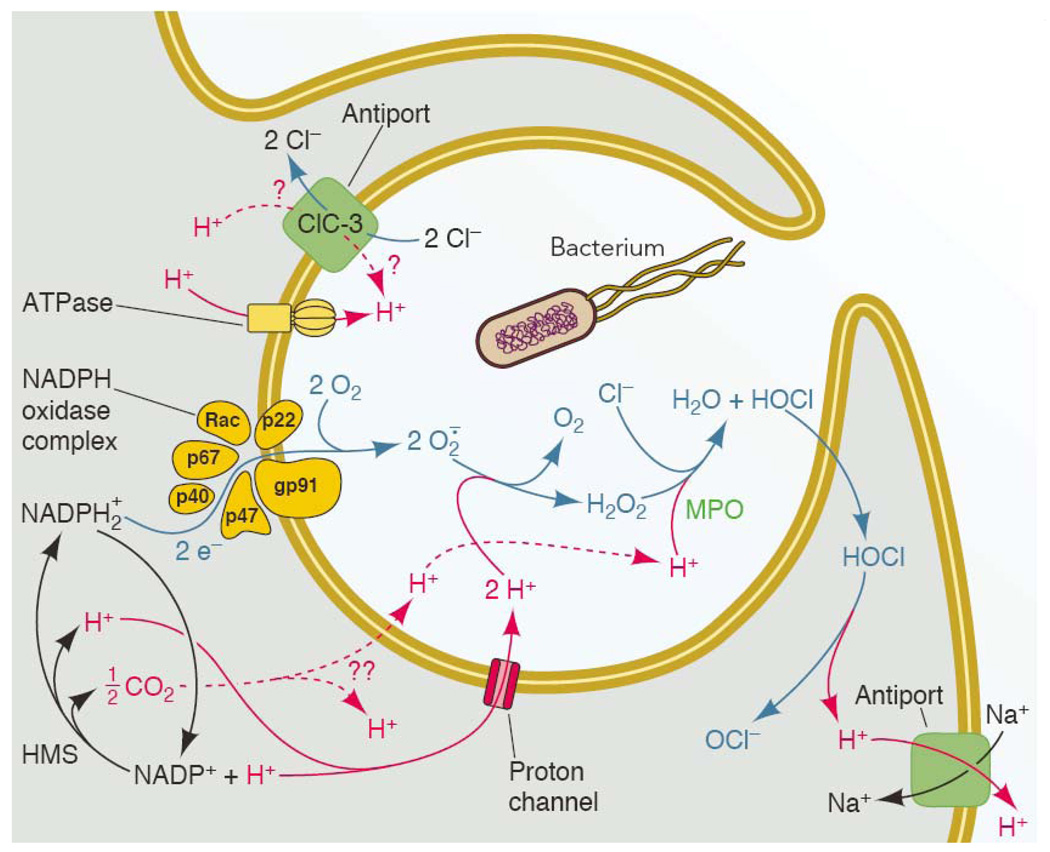
The main molecules and transporters thought to participate in charge compensation and pH regulation during the respiratory burst. A phagocyte is depicted engulfing a bacterium into a nascent phagosome, which will proceed to close and become intracellular. NADPH oxidase assembles preferentially in the phagosomal membrane in neutrophils (19, 167), and begins to function before the phagocytic cup has sealed (119). In eosinophils (111) macrophages (98), and neutrophils stimulated with soluble agonists (16), most NADPH oxidase assembles at the plasma membrane. Charge compensation is required wherever the enzyme is located. The entire system is driven by NADPH oxidase activity. Electrons from cytoplasmic NADPH are translocated across a redox chain to reduce O2 to superoxide anion, O2.−, inside the phagosome or extracellularly. The electrogenic activity can be measured directly as electron current (Fig. 3; 56, 187). For each electron removed from the cell, approximately one proton is left behind. Thus NADPH oxidase activity tends to depolarize the membrane, decrease pHi and increase pHo or pHphagosome. NADPH is regenerated continuously by the hexose monophosphate shunt (HMS) during the respiratory burst. Many of the transporters are functionally unidirectional. An exception is ClC-3 (Table 1), which is shown moving H+ into the phagosome and Cl− out, as is expected to occur at depolarized potentials that exist during the respiratory burst (113). In endosomes lacking Nox activity, ClC-3 is thought to operate in the reverse direction, removing H+ and injecting Cl− into the interior to compensate for electrogenic H+-ATPase activity (96). Although HOCl is membrane permeant (214), it reacts rapidly with cytoplasmic contents such as taurine (129) or glutathione (217); for present purposes, HOCl simply shuttles protons out of the phagosome (135). Note that the H+ in any compartment are equivalent; e.g., protons derived from HOCl dissociation are not preferentially removed by Na+/H+ antiport. [Expanded from ref. (151).]
References
-
- Ädelroth P, Brzezinski P. Surface-mediated proton-transfer reactions in membrane-bound proteins. Biochim Biophys Acta. 2004;1655:102–115. - PubMed
-
- Aggarwal SK, MacKinnon R. Contribution of the S4 segment to gating charge in the Shaker K+ channel. Neuron. 1996;16:1169–1177. - PubMed
-
- Åkerfeldt KS, Lear JD, Wasserman ZR, Chung LA, DeGrado WF. Synthetic peptides as models for ion channel proteins. Acc Chem Res. 1993;26:191–197.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases