

Structure-function study of the N-terminal domain of exocyst subunit Sec3
- PMID: 20139078
- PMCID: PMC2856249
- DOI: 10.1074/jbc.M109.096966
Structure-function study of the N-terminal domain of exocyst subunit Sec3
Abstract
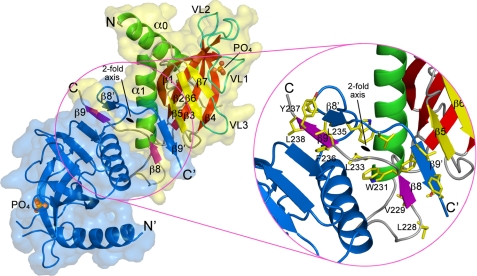
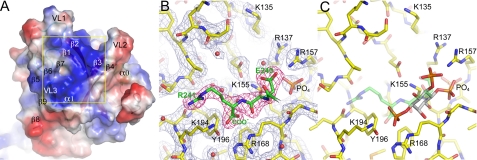
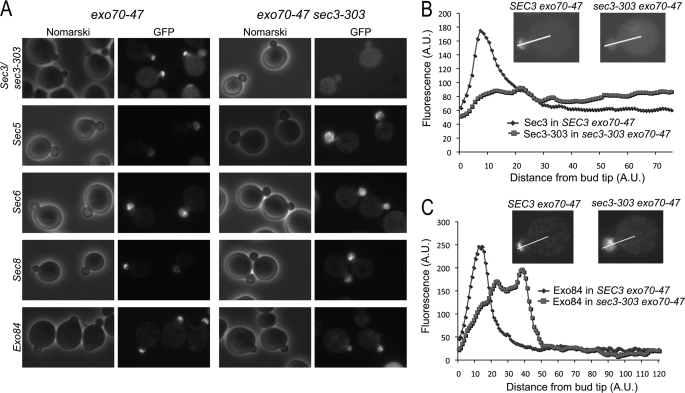
The exocyst is an evolutionarily conserved octameric complex involved in polarized exocytosis from yeast to humans. The Sec3 subunit of the exocyst acts as a spatial landmark for exocytosis through its ability to bind phospholipids and small GTPases. The structure of the N-terminal domain of Sec3 (Sec3N) was determined ab initio and defines a new subclass of pleckstrin homology (PH) domains along with a new family of proteins carrying this domain. Respectively, N- and C-terminal to the PH domain Sec3N presents an additional alpha-helix and two beta-strands that mediate dimerization through domain swapping. The structure identifies residues responsible for phospholipid binding, which when mutated in cells impair the localization of exocyst components at the plasma membrane and lead to defects in exocytosis. Through its ability to bind the small GTPase Cdc42 and phospholipids, the PH domain of Sec3 functions as a coincidence detector at the plasma membrane.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
