

Human and non-human primate genomes share hotspots of positive selection
- PMID: 20140238
- PMCID: PMC2816677
- DOI: 10.1371/journal.pgen.1000840
Human and non-human primate genomes share hotspots of positive selection
Abstract
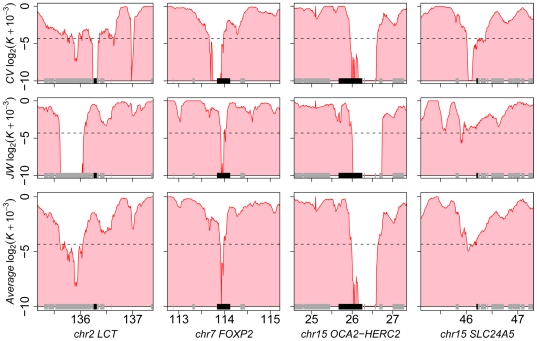
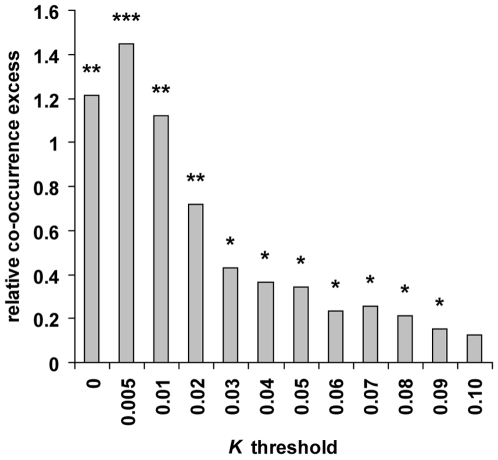
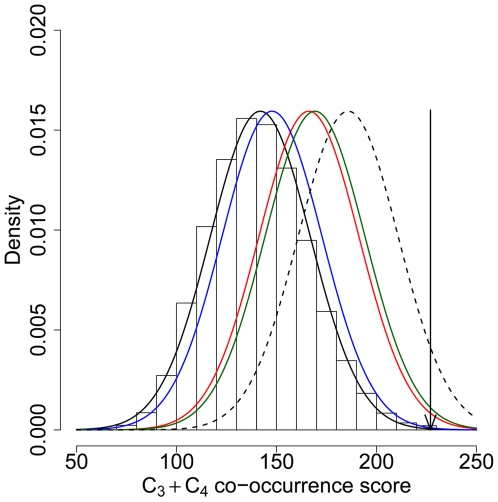
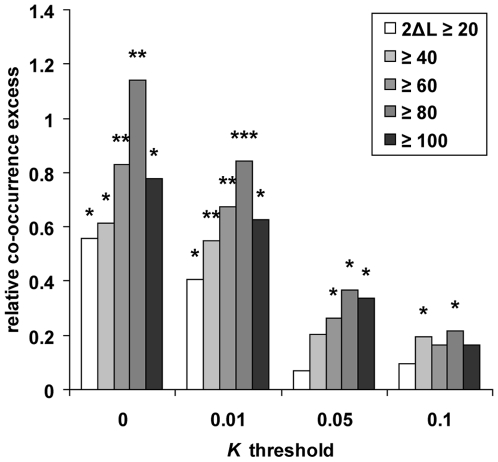
Among primates, genome-wide analysis of recent positive selection is currently limited to the human species because it requires extensive sampling of genotypic data from many individuals. The extent to which genes positively selected in human also present adaptive changes in other primates therefore remains unknown. This question is important because a gene that has been positively selected independently in the human and in other primate lineages may be less likely to be involved in human specific phenotypic changes such as dietary habits or cognitive abilities. To answer this question, we analysed heterozygous Single Nucleotide Polymorphisms (SNPs) in the genomes of single human, chimpanzee, orangutan, and macaque individuals using a new method aiming to identify selective sweeps genome-wide. We found an unexpectedly high number of orthologous genes exhibiting signatures of a selective sweep simultaneously in several primate species, suggesting the presence of hotspots of positive selection. A similar significant excess is evident when comparing genes positively selected during recent human evolution with genes subjected to positive selection in their coding sequence in other primate lineages and identified using a different test. These findings are further supported by comparing several published human genome scans for positive selection with our findings in non-human primate genomes. We thus provide extensive evidence that the co-occurrence of positive selection in humans and in other primates at the same genetic loci can be measured with only four species, an indication that it may be a widespread phenomenon. The identification of positive selection in humans alongside other primates is a powerful tool to outline those genes that were selected uniquely during recent human evolution.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
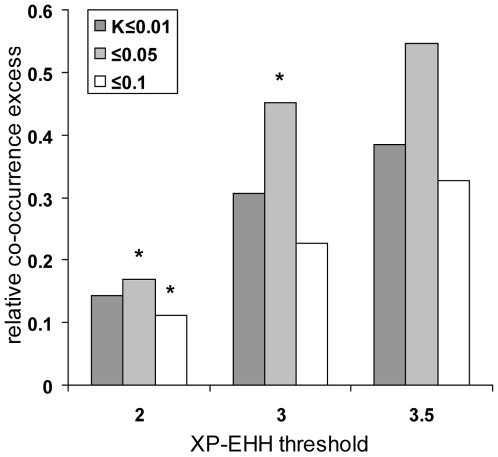
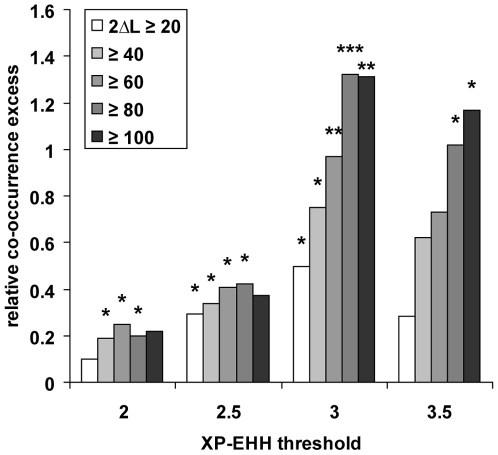
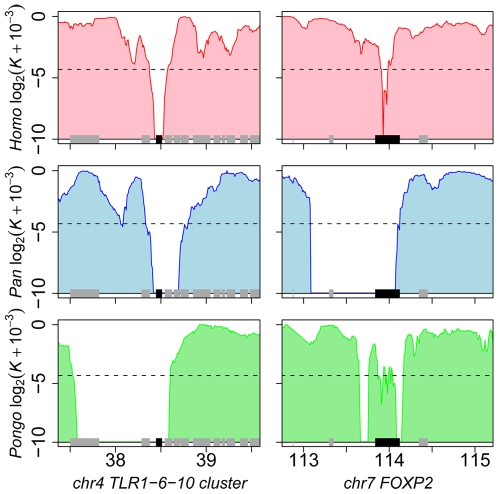
References
-
- Kimura M. The neutral theory of molecular evolution. Sci Am. 1979;241:98–100, 102, 108 passim. - PubMed
-
- Eyre-Walker A, Keightley PD. The distribution of fitness effects of new mutations. Nat Rev Genet. 2007;8:610–618. - PubMed
-
- Boyko AR, Williamson SH, Indap AR, Degenhardt JD, Hernandez RD, et al. Assessing the evolutionary impact of amino acid mutations in the human genome. PLoS Genet. 2008;4:e1000083. doi: 10.1371/journal.pgen.1000083. - DOI - PMC - PubMed
-
- Charlesworth J, Eyre-Walker A. The rate of adaptive evolution in enteric bacteria. Mol Biol Evol. 2006;23:1348–1356. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
