

Review
doi: 10.1002/dvdy.22244.
Integration of diverse inputs in the regulation of Caenorhabditis elegans DAF-16/FOXO
Affiliations
- PMID: 20140911
- PMCID: PMC3811053
- DOI: 10.1002/dvdy.22244
Item in Clipboard
Review
Integration of diverse inputs in the regulation of Caenorhabditis elegans DAF-16/FOXO
Dev Dyn.
2010 May.
Abstract
In a remarkably conserved insulin signaling pathway that is well-known for its regulation of longevity in worms, flies, and mammals, the major C. elegans effector of this pathway, DAF-16/FOXO, also modulates many other physiological processes. This raises the question of how DAF-16/FOXO chooses the correct targets to achieve the appropriate response in a particular context. Here, we review current knowledge of tissue-specificity and interacting partners that modulate DAF-16/FOXO functional output.
Copyright (c) 2010 Wiley-Liss, Inc.
Figures

Activated DAF-2 initiates a phosphorylation cascade that ultimately phosphorylates DAF-16, thereby excluding it from the nucleus and inhibiting its activity. As noted in the text, it remains unclear whether DAF-16 interacts with both 14–3-3 proteins FTT-2 and PAR-5, so here we have labeled its interacting partner “14–3-3.”
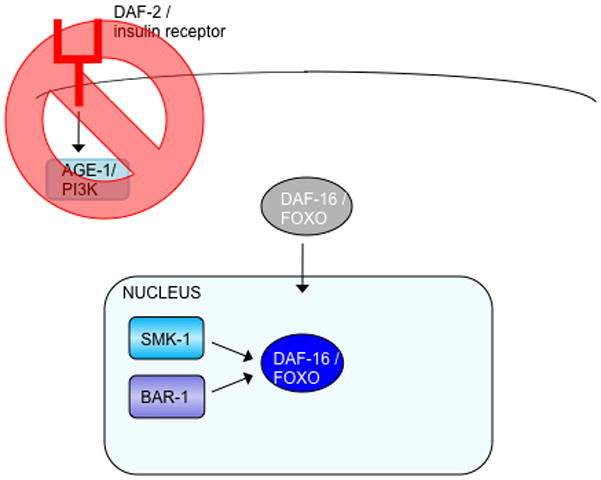
(a) Reduced insulin signaling relieves the phosphorylation of DAF-16, allowing it to enter the nucleus and interact with SMK-1 and BAR-1. (b) In response to oxidative stress, JNK-1 activates DAF-16 by phosphorylation. (Note that the phosphatase that may remove the inhibitory AKT/SGK phosphates to allow nuclear translocation is unknown, but the serine/threonine phosphatase PP2A dephosphorylates mammalian FOXO1 (Yan et al., 2008).) Activated DAF-16 accumulates in the nucleus, where it interacts with SIR-2.1/14–3-3, SMK-1, BAR-1, SKN-1, and HCF-1. (c). Heat shock causes JNK-1 to phosphorylate and activate DAF-16, which in turn, enters the nucleus and interacts with SIR-2.1/14–3-3 and HSF-1.

(a) Reduced insulin signaling relieves the phosphorylation of DAF-16, allowing it to enter the nucleus and interact with SMK-1 and BAR-1. (b) In response to oxidative stress, JNK-1 activates DAF-16 by phosphorylation. (Note that the phosphatase that may remove the inhibitory AKT/SGK phosphates to allow nuclear translocation is unknown, but the serine/threonine phosphatase PP2A dephosphorylates mammalian FOXO1 (Yan et al., 2008).) Activated DAF-16 accumulates in the nucleus, where it interacts with SIR-2.1/14–3-3, SMK-1, BAR-1, SKN-1, and HCF-1. (c). Heat shock causes JNK-1 to phosphorylate and activate DAF-16, which in turn, enters the nucleus and interacts with SIR-2.1/14–3-3 and HSF-1.
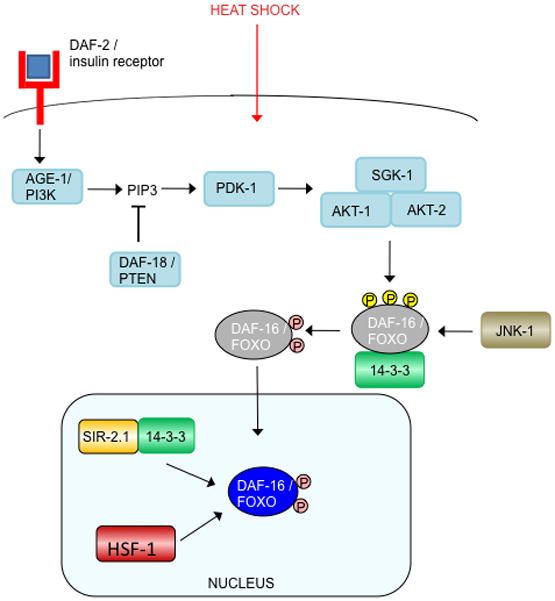
(a) Reduced insulin signaling relieves the phosphorylation of DAF-16, allowing it to enter the nucleus and interact with SMK-1 and BAR-1. (b) In response to oxidative stress, JNK-1 activates DAF-16 by phosphorylation. (Note that the phosphatase that may remove the inhibitory AKT/SGK phosphates to allow nuclear translocation is unknown, but the serine/threonine phosphatase PP2A dephosphorylates mammalian FOXO1 (Yan et al., 2008).) Activated DAF-16 accumulates in the nucleus, where it interacts with SIR-2.1/14–3-3, SMK-1, BAR-1, SKN-1, and HCF-1. (c). Heat shock causes JNK-1 to phosphorylate and activate DAF-16, which in turn, enters the nucleus and interacts with SIR-2.1/14–3-3 and HSF-1.
Similar articles
-
daf-16/FOXO blocks adult cell fate in Caenorhabditis elegans dauer larvae via lin-41/TRIM71.PLoS Genet. 2021 Nov 15;17(11):e1009881. doi: 10.1371/journal.pgen.1009881. eCollection 2021 Nov. PLoS Genet. 2021. PMID: 34780472 Free PMC article.
-
EAK proteins: novel conserved regulators of C. elegans lifespan.Aging (Albany NY). 2010 Oct;2(10):742-7. doi: 10.18632/aging.100214. Aging (Albany NY). 2010. PMID: 20975207 Free PMC article.
-
Longevity Genes Revealed by Integrative Analysis of Isoform-Specific daf-16/FoxO Mutants of Caenorhabditis elegans.Genetics. 2015 Oct;201(2):613-29. doi: 10.1534/genetics.115.177998. Epub 2015 Jul 27. Genetics. 2015. PMID: 26219299 Free PMC article.
-
The search for DAF-16/FOXO transcriptional targets: approaches and discoveries.Exp Gerontol. 2006 Oct;41(10):910-21. doi: 10.1016/j.exger.2006.06.040. Epub 2006 Aug 24. Exp Gerontol. 2006. PMID: 16934425 Review.
-
DAF-16: FOXO in the Context of C. elegans.Curr Top Dev Biol. 2018;127:1-21. doi: 10.1016/bs.ctdb.2017.11.007. Epub 2018 Feb 2. Curr Top Dev Biol. 2018. PMID: 29433733 Review.
Cited by
-
The conserved histone chaperone LIN-53 is required for normal lifespan and maintenance of muscle integrity in Caenorhabditis elegans.Aging Cell. 2019 Dec;18(6):e13012. doi: 10.1111/acel.13012. Epub 2019 Aug 9. Aging Cell. 2019. PMID: 31397537 Free PMC article.
-
The hypoxia-inducible factor HIF-1 functions as both a positive and negative modulator of aging.Biol Chem. 2010 Oct;391(10):1131-7. doi: 10.1515/BC.2010.123. Biol Chem. 2010. PMID: 20707608 Free PMC article. Review.
-
The neurodegenerative effects of selenium are inhibited by FOXO and PINK1/PTEN regulation of insulin/insulin-like growth factor signaling in Caenorhabditis elegans.Neurotoxicology. 2014 Mar;41(100):28-43. doi: 10.1016/j.neuro.2013.12.012. Epub 2014 Jan 6. Neurotoxicology. 2014. PMID: 24406377 Free PMC article.
-
Folic acid supplementation at lower doses increases oxidative stress resistance and longevity in Caenorhabditis elegans.Age (Dordr). 2015 Dec;37(6):113. doi: 10.1007/s11357-015-9850-5. Epub 2015 Nov 6. Age (Dordr). 2015. PMID: 26546011 Free PMC article.
-
Insulin/FOXO signaling regulates ovarian prostaglandins critical for reproduction.Dev Cell. 2010 Dec 14;19(6):858-71. doi: 10.1016/j.devcel.2010.11.005. Dev Cell. 2010. PMID: 21145501 Free PMC article.
References
-
- Apfeld J, Kenyon C. Cell nonautonomy of C. elegans daf-2 function in the regulation of diapause and life span. Cell. 1998;95:199–210. - PubMed
-
- Baugh LR, Sternberg PW. DAF-16/FOXO regulates transcription of cki-1/Cip/Kip and repression of lin-4 during C. elegans L1 arrest. Curr Biol. 2006;16:780–785. - PubMed
-
- Berdichevsky A, Viswanathan M, Horvitz HR, Guarente L. C. elegans SIR-2.1 interacts with 14–3-3 proteins to activate DAF-16 and extend life span. Cell. 2006;125:1165–1177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
