

Chemical biology strategy reveals pathway-selective inhibitor of NF-kappaB activation induced by protein kinase C
- PMID: 20141195
- PMCID: PMC2842467
- DOI: 10.1021/cb9003089
Chemical biology strategy reveals pathway-selective inhibitor of NF-kappaB activation induced by protein kinase C
Abstract
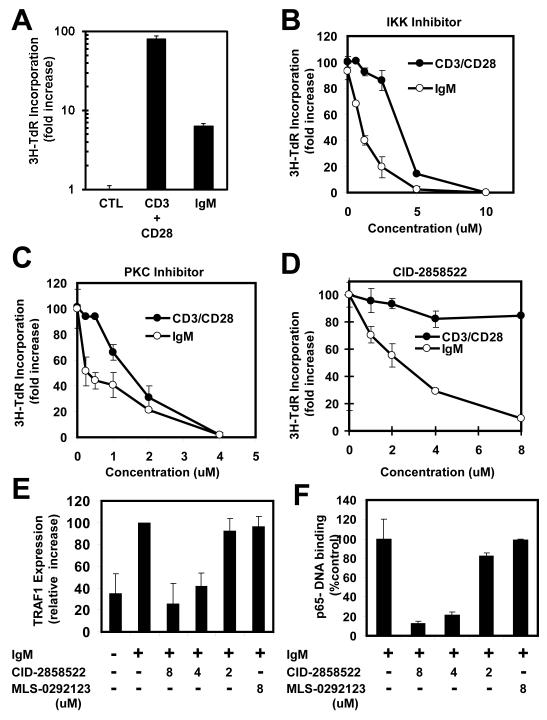
Dysregulation of NF-kappaB activity contributes to many autoimmune and inflammatory diseases. At least nine pathways for NF-kappaB activation have been identified, most of which converge on the IkappaB kinases (IKKs). Although IKKs represent logical targets for potential drug discovery, chemical inhibitors of IKKs suppress all known NF-kappaB activation pathways and thus lack the selectivity required for safe use. A unique NF-kappaB activation pathway is initiated by protein kinase C (PKC) that is stimulated by antigen receptors and many growth factor receptors. Using a cell-based high-throughput screening (HTS) assay and chemical biology strategy, we identified a 2-aminobenzimidazole compound, CID-2858522, which selectively inhibits the NF-kappaB pathway induced by PKC, operating downstream of PKC but upstream of IKKbeta, without inhibiting other NF-kappaB activation pathways. In human B cells stimulated through surface immunoglobulin, CID-2858522 inhibited NF-kappaB DNA-binding activity and expression of endogenous NF-kappaB-dependent target gene, TRAF1. Altogether, as a selective chemical inhibitor of the NF-kappaB pathway induced by PKC, CID-2858522 serves as a powerful research tool and may reveal new paths toward therapeutically useful NF-kappaB inhibitors.
Figures




Similar articles
-
Novel non-peptide small molecules preventing IKKβ/NEMO association inhibit NF-κB activation in LPS-stimulated J774 macrophages.Biochem Pharmacol. 2016 Mar 15;104:83-94. doi: 10.1016/j.bcp.2016.01.008. Epub 2016 Jan 14. Biochem Pharmacol. 2016. PMID: 26776306
-
Protein kinase C-theta participates in NF-kappaB activation induced by CD3-CD28 costimulation through selective activation of IkappaB kinase beta.Mol Cell Biol. 2000 Apr;20(8):2933-40. doi: 10.1128/MCB.20.8.2933-2940.2000. Mol Cell Biol. 2000. PMID: 10733597 Free PMC article.
-
The protein phosphatase 2A regulatory subunit B56γ mediates suppression of T cell receptor (TCR)-induced nuclear factor-κB (NF-κB) activity.J Biol Chem. 2014 May 23;289(21):14996-5004. doi: 10.1074/jbc.M113.533547. Epub 2014 Apr 9. J Biol Chem. 2014. PMID: 24719332 Free PMC article.
-
NF-κB-IKKβ Pathway as a Target for Drug Development: Realities, Challenges and Perspectives.Curr Drug Targets. 2018;19(16):1933-1942. doi: 10.2174/1389450119666180219120534. Curr Drug Targets. 2018. PMID: 29468963 Review.
-
Regulation and function of IKK and IKK-related kinases.Sci STKE. 2006 Oct 17;2006(357):re13. doi: 10.1126/stke.3572006re13. Sci STKE. 2006. PMID: 17047224 Review.
Cited by
-
Inhibition of proliferation and survival of diffuse large B-cell lymphoma cells by a small-molecule inhibitor of the ubiquitin-conjugating enzyme Ubc13-Uev1A.Blood. 2012 Aug 23;120(8):1668-77. doi: 10.1182/blood-2012-02-406074. Epub 2012 Jul 12. Blood. 2012. PMID: 22791293 Free PMC article.
-
Synthesis and physicochemical characterization of novel phenotypic probes targeting the nuclear factor-kappa B signaling pathway.Beilstein J Org Chem. 2013 May 8;9:900-7. doi: 10.3762/bjoc.9.103. Print 2013. Beilstein J Org Chem. 2013. PMID: 23766805 Free PMC article.
-
Trimethylamine N-oxide in atherogenesis: impairing endothelial self-repair capacity and enhancing monocyte adhesion.Biosci Rep. 2017 Mar 2;37(2):BSR20160244. doi: 10.1042/BSR20160244. Print 2017 Apr 30. Biosci Rep. 2017. PMID: 28153917 Free PMC article.
-
Network and Experimental Pharmacology on Mechanism of Yixintai Regulates the TMAO/PKC/NF-κB Signaling Pathway in Treating Heart Failure.Drug Des Devel Ther. 2024 Apr 30;18:1415-1438. doi: 10.2147/DDDT.S448140. eCollection 2024. Drug Des Devel Ther. 2024. PMID: 38707614 Free PMC article.
-
PKC signaling contributes to chromatin decondensation and is required for competence to respond to IL-2 during T cell activation.Cell Immunol. 2020 Jan;347:104027. doi: 10.1016/j.cellimm.2019.104027. Epub 2019 Dec 11. Cell Immunol. 2020. PMID: 31864664 Free PMC article.
References
-
- Tracey KJ, Cerami A. Tumor necrosis factor, other cytokines and disease. Annu Rev Cell Biol. 1993;9:317–43. - PubMed
-
- Karin M, Delhase M. The I kappa B kinase (IKK) and NF-kappa B: key elements of proinflammatory signalling. Semin Immunol. 2000;12(1):85–98. - PubMed
-
- West AP, Koblansky AA, Ghosh S. Recognition and signaling by toll-like receptors. Annu Rev Cell Dev Biol. 2006;22:409–37. - PubMed
-
- Fujita T. Virology. Sensing viral RNA amid your own. Science. 2006;314(5801):935–6. - PubMed
-
- Janssens S, Tschopp J. Signals from within: the DNA-damage-induced NF-kappaB response. Cell Death Differ. 2006;13(5):773–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
