

Genome-wide analysis of rice ClpB/HSP100, ClpC and ClpD genes
- PMID: 20141629
- PMCID: PMC2829514
- DOI: 10.1186/1471-2164-11-95
Genome-wide analysis of rice ClpB/HSP100, ClpC and ClpD genes
Abstract
Background: ClpB-cyt/HSP100 protein acts as chaperone, mediating disaggregation of denatured proteins. Previous studies have shown that ClpB-cyt/HSP100 gene belongs to the group class I Clp ATPase proteins and ClpB-cyt/HSP100 transcript is regulated by heat stress and developmental cues.
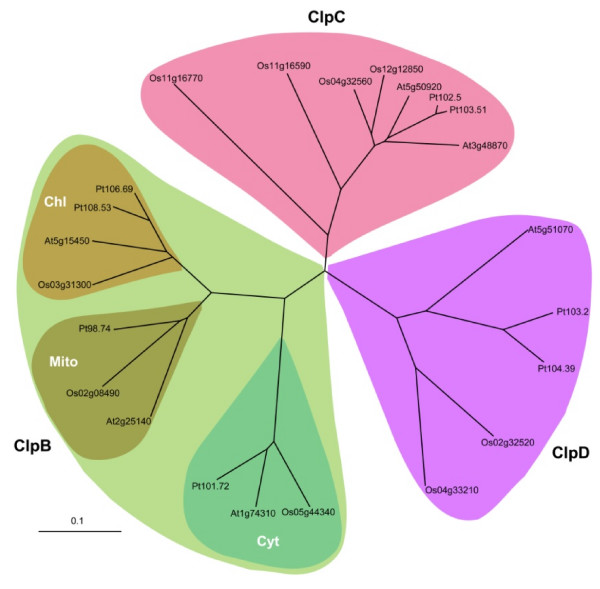
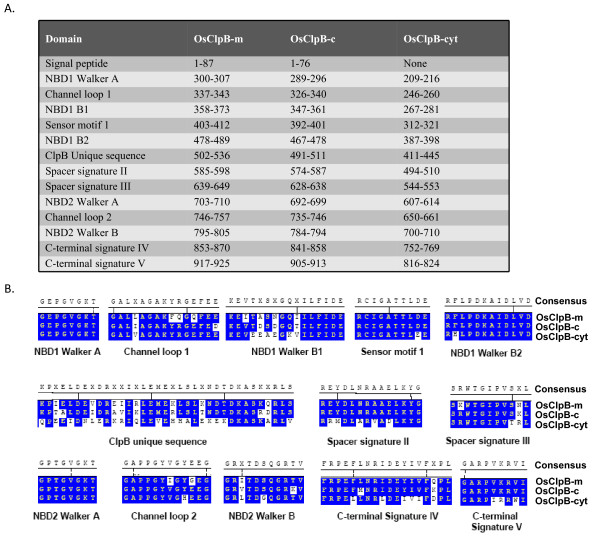
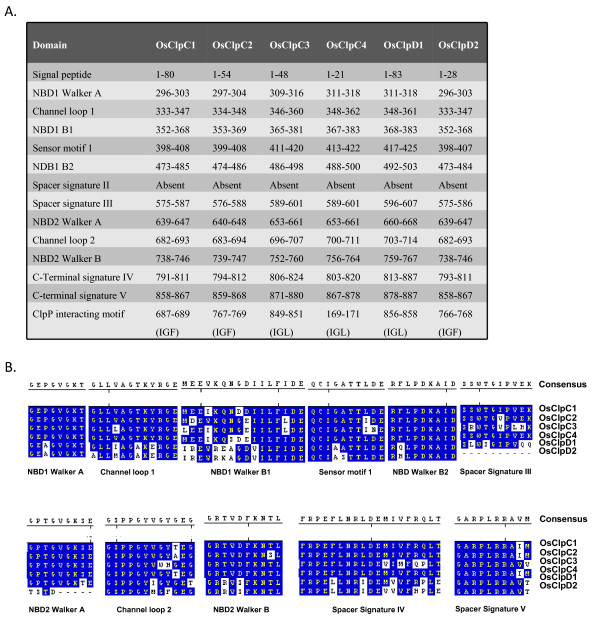
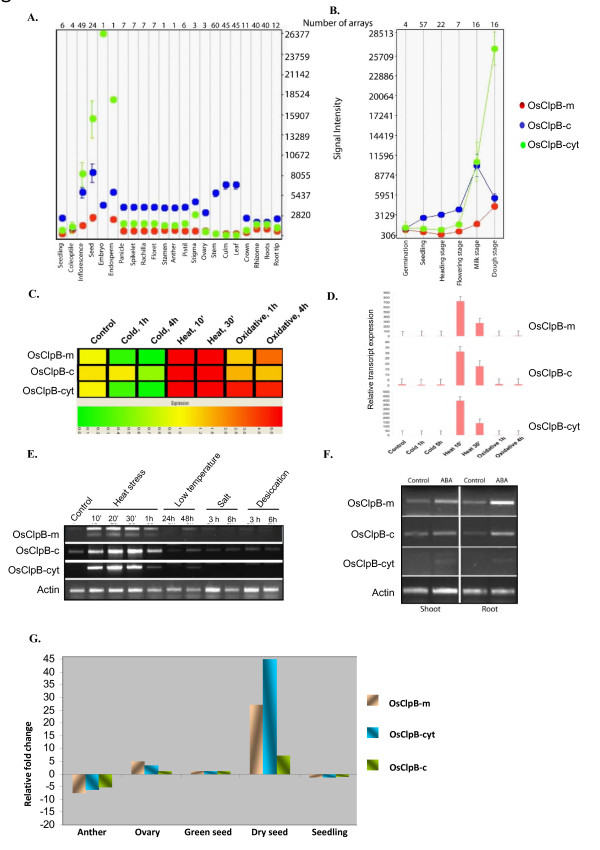
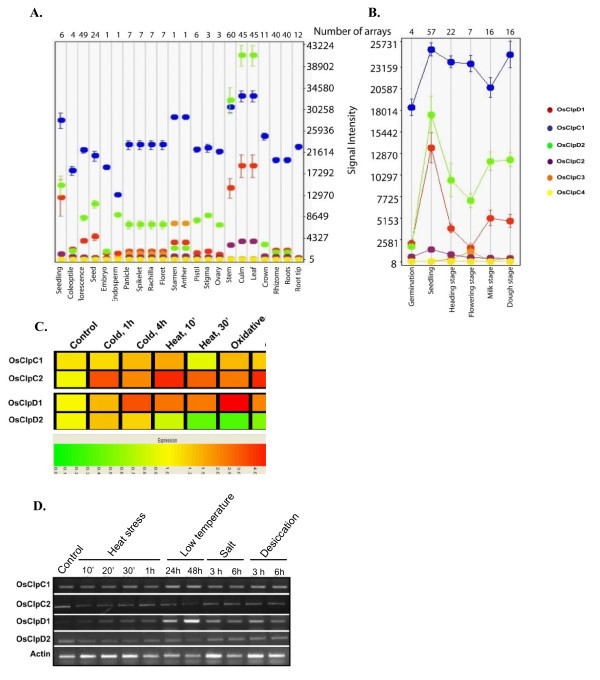
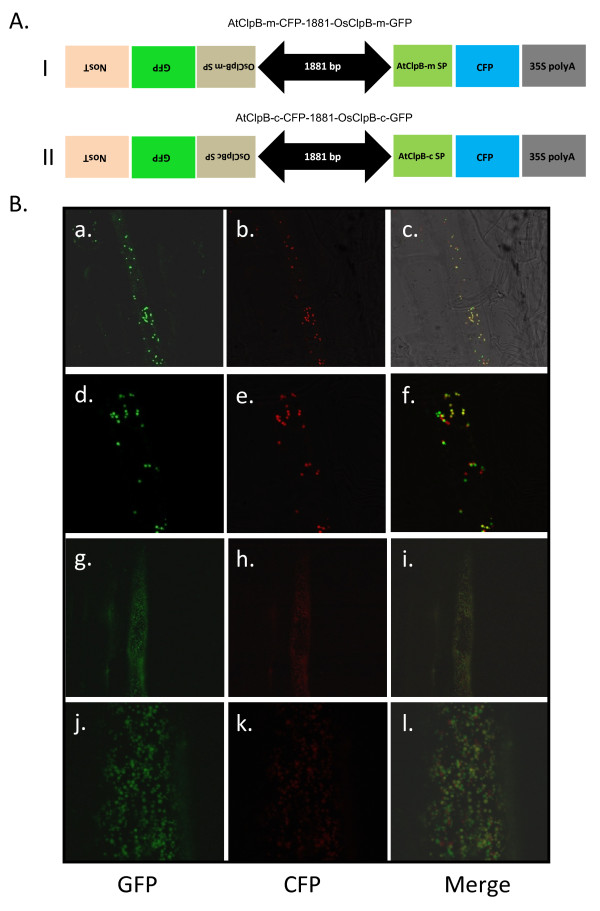
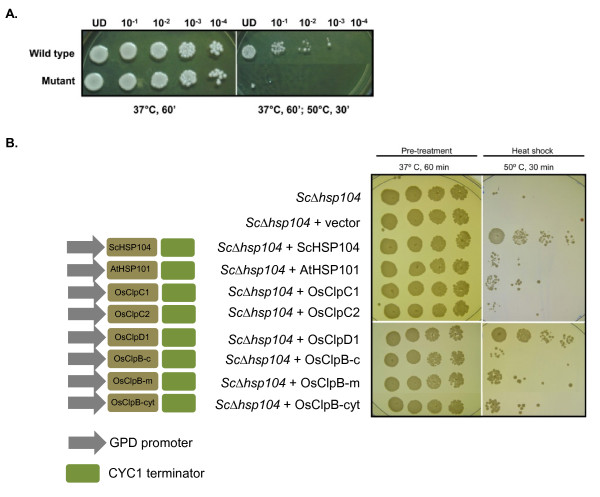
Results: Nine ORFs were noted to constitute rice class I Clp ATPases in the following manner: 3 ClpB proteins (ClpB-cyt, Os05g44340; ClpB-m, Os02g08490; ClpB-c, Os03g31300), 4 ClpC proteins (ClpC1, Os04g32560; ClpC2, Os12g12580; ClpC3, Os11g16590; ClpC4, Os11g16770) and 2 ClpD proteins (ClpD1, Os02g32520; ClpD2, Os04g33210). Using the respective signal sequences cloned upstream to GFP/CFP reporter proteins and transient expression studies with onion epidermal cells, evidence is provided that rice ClpB-m and Clp-c proteins are indeed localized to their respective cell locations mitochondria and chloroplasts, respectively. Associated with their diverse cell locations, domain structures of OsClpB-c, OsClpB-m and OsClpB-cyt proteins are noted to possess a high-level conservation. OsClpB-cyt transcript is shown to be enriched at milk and dough stages of seed development. While expression of OsClpB-m was significantly less as compared to its cytoplasmic and chloroplastic counterparts in different tissues, this transcript showed highest heat-induced expression amongst the 3 ClpB proteins. OsClpC1 and OsClpC2 are predicted to be chloroplast-localized as is the case with all known plant ClpC proteins. However, the fact that OsClpC3 protein appears mitochondrial/chloroplastic with equal probability and OsClpC4 a plasma membrane protein reflects functional diversity of this class. Different class I Clp ATPase transcripts were noted to be cross-induced by a host of different abiotic stress conditions. Complementation assays of Deltahsp104 mutant yeast cells showed that OsClpB-cyt, OsClpB-m, OsClpC1 and OsClpD1 have significantly positive effects. Remarkably, OsClpD1 gene imparted appreciably high level tolerance to the mutant yeast cells.
Conclusions: Rice class I Clp ATPase gene family is constituted of 9 members. Of these 9, only 3 belonging to ClpB group are heat stress regulated. Distribution of ClpB proteins to different cell organelles indicates that their functioning might be critical in different cell locations. From the complementation assays, OsClpD1 appears to be more effective than OsClpB-cyt protein in rescuing the thermosensitive defect of the yeast ScDeltahsp104 mutant cells.
Figures
References
-
- Grover A, Chandramouli A, Agarwal S, Katiyar-Agarwal S, Agarwal M, Sahi C. In: Rice Improvement in the Genomic Era. Dutta SK, editor. Hawarth Press USA; 2009. Transgenic rice for tolerance against abiotic stresses; pp. 237–267.
-
- Frank G, Pressman E, Ophir R, Althan L, Shaked R, Freedman M, Shen S, Firon N. Transcriptional profiling of maturing tomato (Solanum lycopersicum L.) microspores reveals the involvement of heat shock proteins, ROS scavengers, hormones, and sugars in the heat stress response. Journal of Experimental Botany. 2009;60(13):3891–3908. doi: 10.1093/jxb/erp234. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
