

Complementary interplay between matrix metalloproteinase-9, vascular endothelial growth factor and osteoclast function drives endochondral bone formation
- PMID: 20142327
- PMCID: PMC2869494
- DOI: 10.1242/dmm.004226
Complementary interplay between matrix metalloproteinase-9, vascular endothelial growth factor and osteoclast function drives endochondral bone formation
Abstract
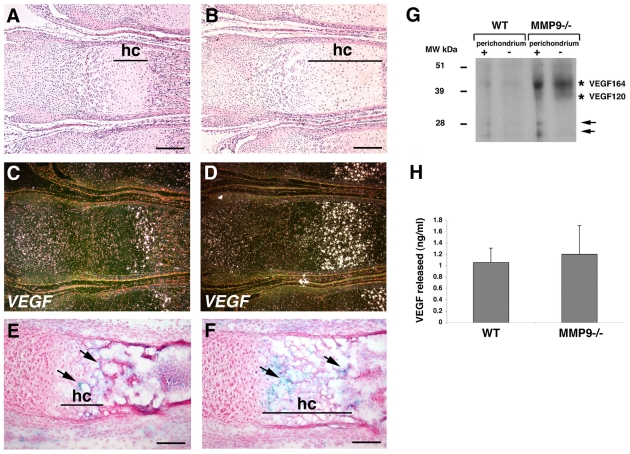
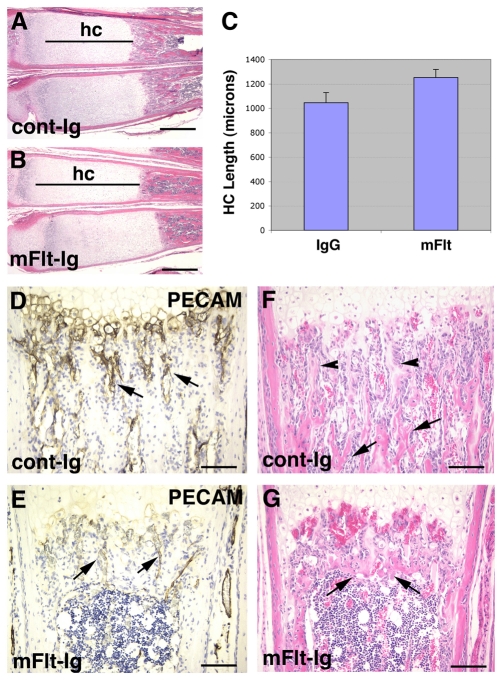
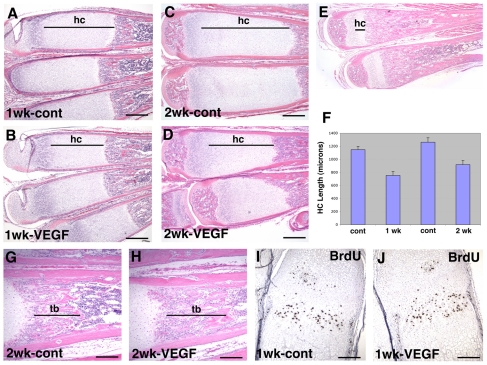
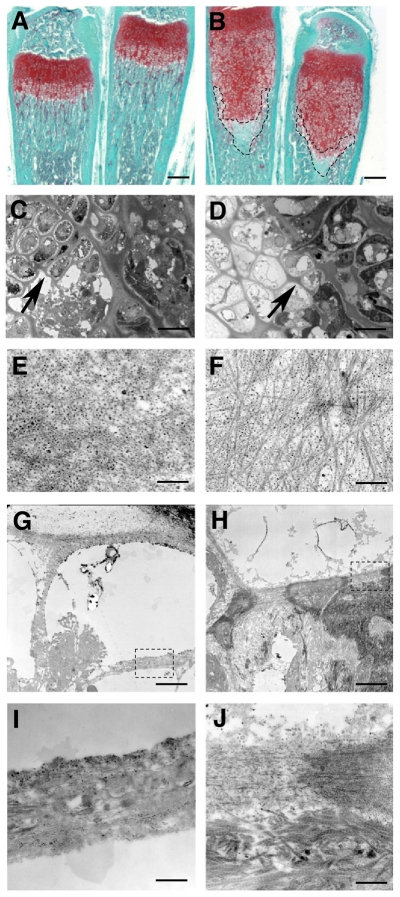
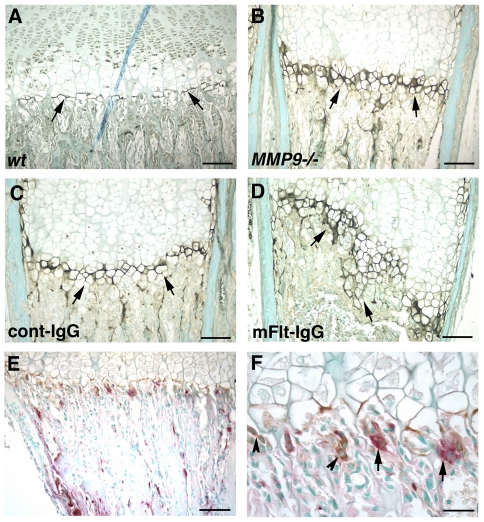
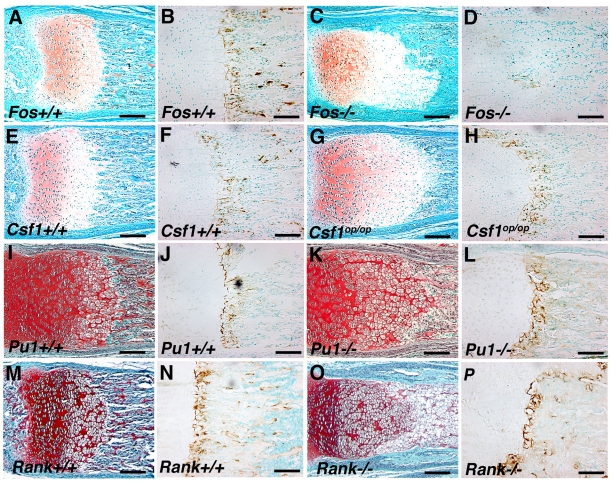
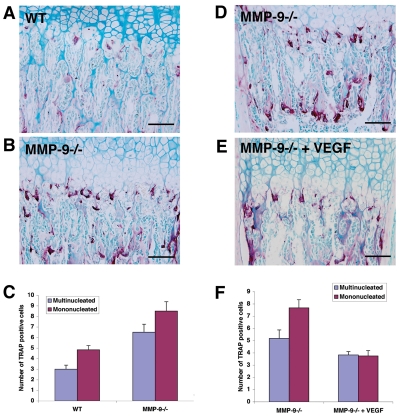
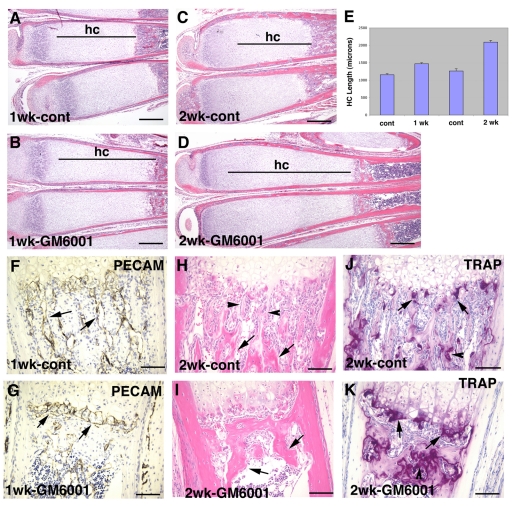
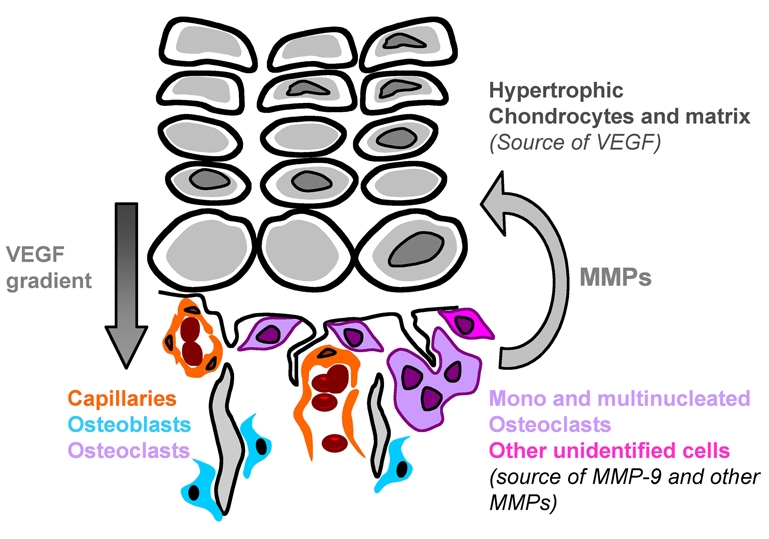
Long bone development depends on endochondral bone formation, a complex process requiring exquisite balance between hypertrophic cartilage (HC) formation and its ossification. Dysregulation of this process may result in skeletal dysplasias and heterotopic ossification. Endochondral ossification requires the precise orchestration of HC vascularization, extracellular matrix remodeling, and the recruitment of osteoclasts and osteoblasts. Matrix metalloproteinase-9 (MMP-9), vascular endothelial growth factor (VEGF) and osteoclasts have all been shown to regulate endochondral ossification, but how their function interrelates is not known. We have investigated the functional relationship among these regulators of endochondral ossification, demonstrating that they have complementary but non-overlapping functions. MMP-9, VEGF and osteoclast deficiency all cause impaired growth plate ossification resulting in the accumulation of HC. VEGF mRNA and protein expression are increased at the MMP-9-/- growth plate, and VEGF activity contributes to endochondral ossification since sequestration of VEGF by soluble receptors results in further inhibition of growth plate vascularization and ossification. However, VEGF bioavailability is still limited in MMP-9 deficiency, as exogenous VEGF is able to rescue the MMP-9-/- phenotype, demonstrating that MMP-9 may partially, but not fully, regulate VEGF bioavailability. The organization of the HC extracellular matrix at the MMP-9-/- growth plate is altered, supporting a role for MMP-9 in HC remodeling. Inhibition of VEGF impairs osteoclast recruitment, whereas MMP-9 deficiency leads to an accumulation of osteoclasts at the chondro-osseous junction. Growth plate ossification in osteoclast-deficient mice is impaired in the presence of normal MMP-9 expression, indicating that other osteoclastic functions are also necessary. Our data delineate the complementary interplay between MMP-9, VEGF and osteoclast function that is necessary for normal endochondral bone formation and provide a molecular framework for investigating the molecular defects contributing to disorders of endochondral bone formation.
Figures

Similar articles
-
The critical role of the epidermal growth factor receptor in endochondral ossification.J Bone Miner Res. 2011 Nov;26(11):2622-33. doi: 10.1002/jbmr.502. J Bone Miner Res. 2011. PMID: 21887704 Free PMC article.
-
Perlecan modulates VEGF signaling and is essential for vascularization in endochondral bone formation.Matrix Biol. 2012 May;31(4):234-45. doi: 10.1016/j.matbio.2012.02.006. Epub 2012 Mar 7. Matrix Biol. 2012. PMID: 22421594 Free PMC article.
-
Chondrocytic ephrin B2 promotes cartilage destruction by osteoclasts in endochondral ossification.Development. 2016 Feb 15;143(4):648-57. doi: 10.1242/dev.125625. Epub 2016 Jan 11. Development. 2016. PMID: 26755702
-
The skeleton: a multi-functional complex organ: the growth plate chondrocyte and endochondral ossification.J Endocrinol. 2011 Nov;211(2):109-21. doi: 10.1530/JOE-11-0048. Epub 2011 Jun 3. J Endocrinol. 2011. PMID: 21642379 Review.
-
The hypertrophic chondrocyte: To be or not to be.Histol Histopathol. 2021 Oct;36(10):1021-1036. doi: 10.14670/HH-18-355. Epub 2021 Jun 17. Histol Histopathol. 2021. PMID: 34137454 Free PMC article. Review.
Cited by
-
c-Raf promotes angiogenesis during normal growth plate maturation.Development. 2016 Jan 15;143(2):348-55. doi: 10.1242/dev.127142. Epub 2015 Dec 10. Development. 2016. PMID: 26657770 Free PMC article.
-
Epidermal growth factor receptor (EGFR) signaling regulates epiphyseal cartilage development through β-catenin-dependent and -independent pathways.J Biol Chem. 2013 Nov 8;288(45):32229-32240. doi: 10.1074/jbc.M113.463554. Epub 2013 Sep 18. J Biol Chem. 2013. PMID: 24047892 Free PMC article.
-
Choline kinase beta is required for normal endochondral bone formation.Biochim Biophys Acta. 2014 Jul;1840(7):2112-22. doi: 10.1016/j.bbagen.2014.03.008. Epub 2014 Mar 14. Biochim Biophys Acta. 2014. PMID: 24637075 Free PMC article.
-
Lysophosphatidic acid enhanced the angiogenic capability of human chondrocytes by regulating Gi/NF-kB-dependent angiogenic factor expression.PLoS One. 2014 May 30;9(5):e95180. doi: 10.1371/journal.pone.0095180. eCollection 2014. PLoS One. 2014. PMID: 24879414 Free PMC article.
-
The traumatic bone: trauma-induced heterotopic ossification.Transl Res. 2017 Aug;186:95-111. doi: 10.1016/j.trsl.2017.06.004. Epub 2017 Jun 15. Transl Res. 2017. PMID: 28668522 Free PMC article. Review.
References
-
- Aldridge SE, Lennard TW, Williams JR, Birch MA. (2005). Vascular endothelial growth factor receptors in osteoclast differentiation and function. Biochem Biophys Res Commun. 335, 793–798 - PubMed
-
- Arikawa-Hirasawa E, Watanabe H, Takami H, Hassell JR, Yamada Y. (1999). Perlecan is essential for cartilage and cephalic development. Nat Genet. 23, 354–358 - PubMed
-
- Bischof P, Truong K, Campana A. (2003). Regulation of trophoblastic gelatinases by proto-oncogenes. Placenta 24, 155–163 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
