

The psychedelic genes of maize redundantly promote carbohydrate export from leaves
- PMID: 20142436
- PMCID: PMC2870957
- DOI: 10.1534/genetics.109.113357
The psychedelic genes of maize redundantly promote carbohydrate export from leaves
Abstract
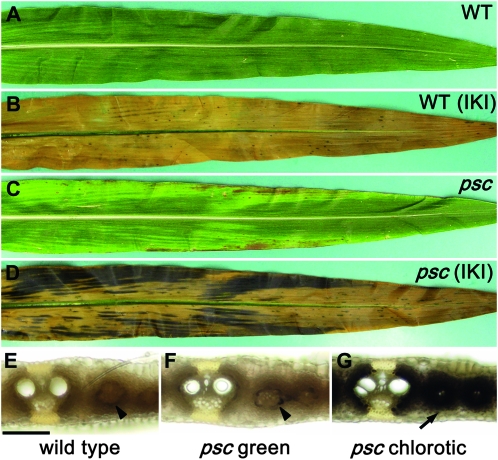
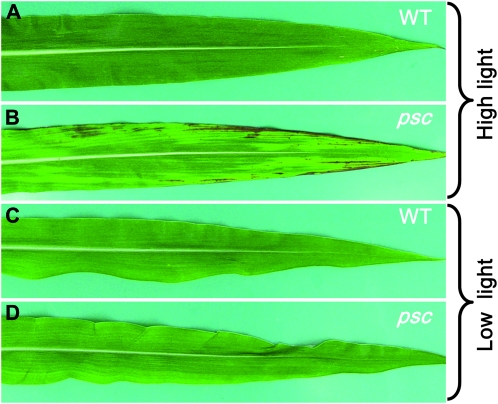
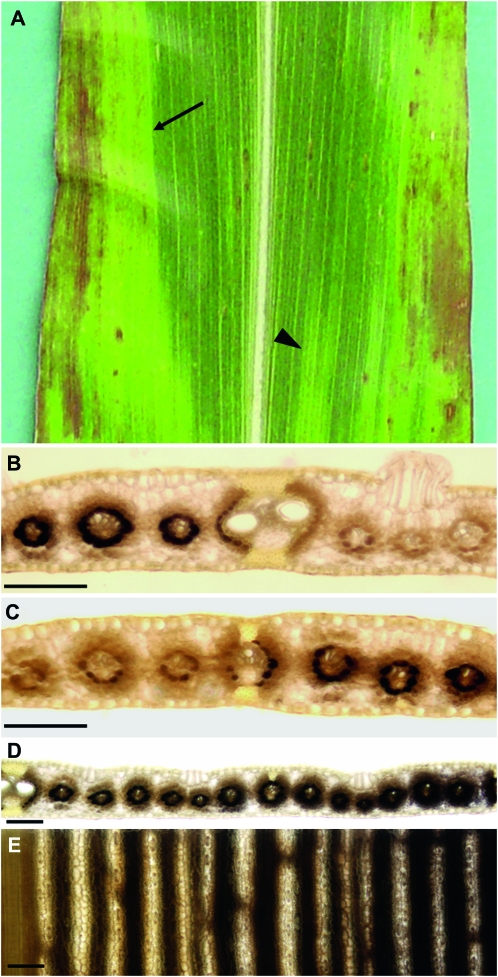
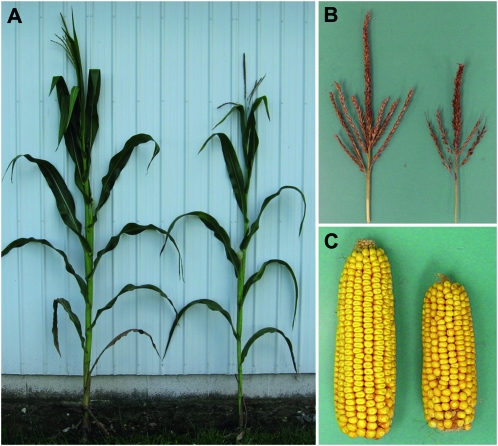
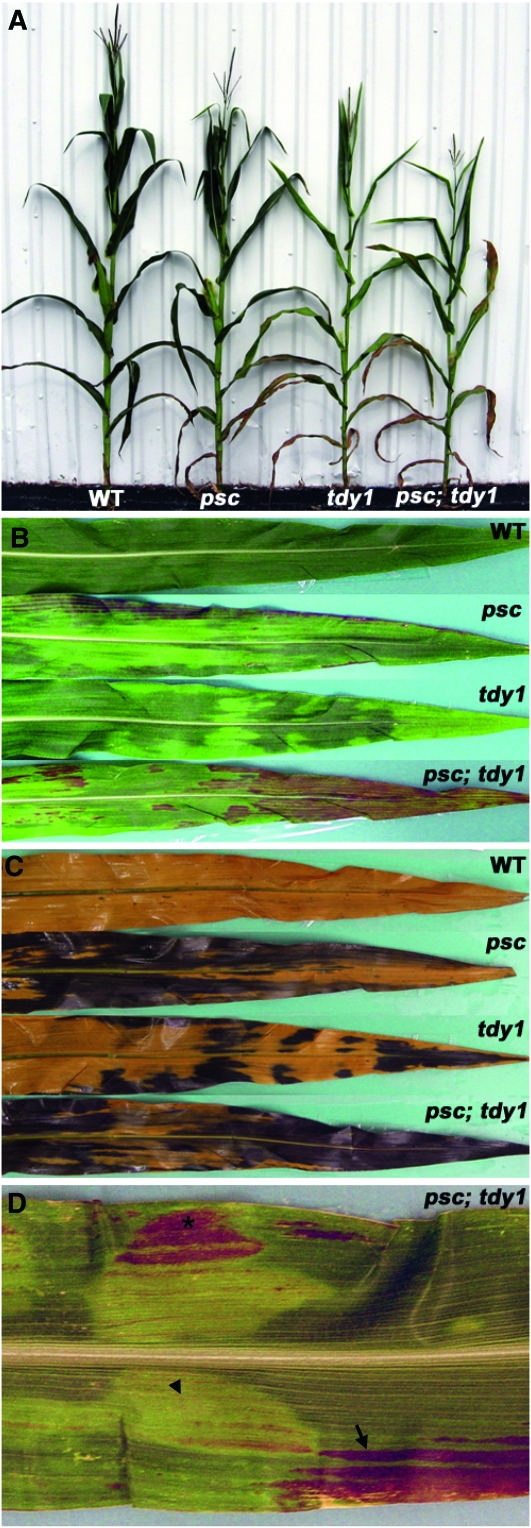
Whole-plant carbohydrate partitioning involves the assimilation of carbon in leaves and its translocation to nonphotosynthetic tissues. This process is fundamental to plant growth and development, but its regulation is poorly understood. To identify genes controlling carbohydrate partitioning, we isolated mutants that are defective in exporting fixed carbon from leaves. Here we describe psychedelic (psc), a new mutant of maize (Zea mays) that is perturbed in carbohydrate partitioning. psc mutants exhibit stable, discrete chlorotic and green regions within their leaves. psc chlorotic tissues hyperaccumulate starch and soluble sugars, while psc green tissues appear comparable to wild-type leaves. The psc chlorotic and green tissue boundaries are usually delineated by larger veins, suggesting that translocation of a mobile compound through the veins may influence the tissue phenotype. psc mutants display altered biomass partitioning, which is consistent with reduced carbohydrate export from leaves to developing tissues. We determined that the psc mutation is unlinked to previously characterized maize leaf carbohydrate hyperaccumulation mutants. Additionally, we found that the psc mutant phenotype is inherited as a recessive, duplicate-factor trait in some inbred lines. Genetic analyses with other maize mutants with variegated leaves and impaired carbohydrate partitioning suggest that Psc defines an independent pathway. Therefore, investigations into the psc mutation have uncovered two previously unknown genes that redundantly function to regulate carbohydrate partitioning in maize.
Figures


Similar articles
-
Tie-dyed2 functions with tie-dyed1 to promote carbohydrate export from maize leaves.Plant Physiol. 2008 Mar;146(3):1085-97. doi: 10.1104/pp.107.111476. Epub 2008 Jan 24. Plant Physiol. 2008. PMID: 18218972 Free PMC article.
-
tie-dyed1 Regulates carbohydrate accumulation in maize leaves.Plant Physiol. 2006 Dec;142(4):1511-22. doi: 10.1104/pp.106.090381. Epub 2006 Oct 27. Plant Physiol. 2006. PMID: 17071639 Free PMC article.
-
Starch phosphorylase 2 is essential for cellular carbohydrate partitioning in maize.J Integr Plant Biol. 2022 Sep;64(9):1755-1769. doi: 10.1111/jipb.13328. Epub 2022 Aug 23. J Integr Plant Biol. 2022. PMID: 35796344
-
Tie-dyed1 and sucrose export defective1 act independently to promote carbohydrate export from maize leaves.Planta. 2008 Feb;227(3):527-38. doi: 10.1007/s00425-007-0636-6. Epub 2007 Oct 9. Planta. 2008. PMID: 17924136 Free PMC article.
-
The power of classic maize mutants: Driving forward our fundamental understanding of plants.Plant Cell. 2022 Jul 4;34(7):2505-2517. doi: 10.1093/plcell/koac081. Plant Cell. 2022. PMID: 35274692 Free PMC article. Review.
Cited by
-
Plant Productivity and Leaf Starch During Grain Fill Is Linked to QTL Containing Flowering Locus T1 (FT1) in Wheat (Triticum aestivum L.).Plants (Basel). 2025 Feb 7;14(4):512. doi: 10.3390/plants14040512. Plants (Basel). 2025. PMID: 40006771 Free PMC article.
-
QTL analysis for carbon assimilate translocation-related traits during maturity in rice (Oryza sativa L.).Breed Sci. 2019 Jun;69(2):289-296. doi: 10.1270/jsbbs.18203. Epub 2019 Jun 5. Breed Sci. 2019. PMID: 31481838 Free PMC article.
-
A Plate Growth Assay to Quantify Embryonic Root Development of Zea mays.Bio Protoc. 2023 Oct 20;13(20):e4858. doi: 10.21769/BioProtoc.4858. eCollection 2023 Oct 20. Bio Protoc. 2023. PMID: 37900110 Free PMC article.
-
The molecular cloning and clarification of a photorespiratory mutant, oscdm1, using enhancer trapping.Front Genet. 2015 Jul 3;6:226. doi: 10.3389/fgene.2015.00226. eCollection 2015. Front Genet. 2015. PMID: 26191072 Free PMC article.
-
Sucrose Transporter ZmSut1 Expression and Localization Uncover New Insights into Sucrose Phloem Loading.Plant Physiol. 2016 Nov;172(3):1876-1898. doi: 10.1104/pp.16.00884. Epub 2016 Sep 12. Plant Physiol. 2016. PMID: 27621426 Free PMC article.
References
-
- Birchler, J., 1994. Dosage analysis using B-A translocations, pp. 328–329 in The Maize Handbook, edited by M. Freeling and V. Walbot. Springer-Verlag, New York.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
