

Uridylation of mature miRNAs and siRNAs by the MUT68 nucleotidyltransferase promotes their degradation in Chlamydomonas
- PMID: 20142471
- PMCID: PMC2840426
- DOI: 10.1073/pnas.0912632107
Uridylation of mature miRNAs and siRNAs by the MUT68 nucleotidyltransferase promotes their degradation in Chlamydomonas
Abstract
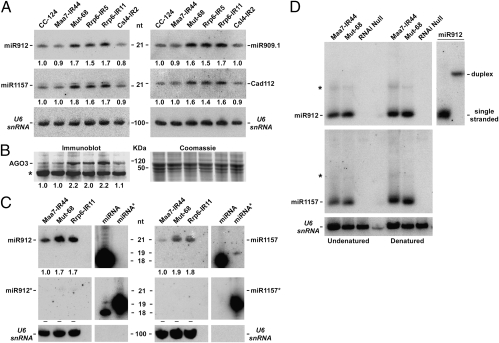
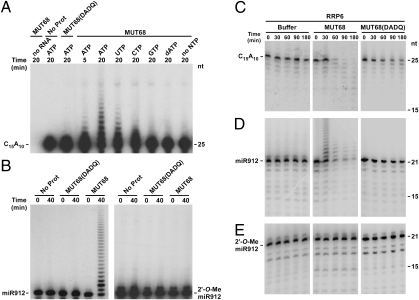
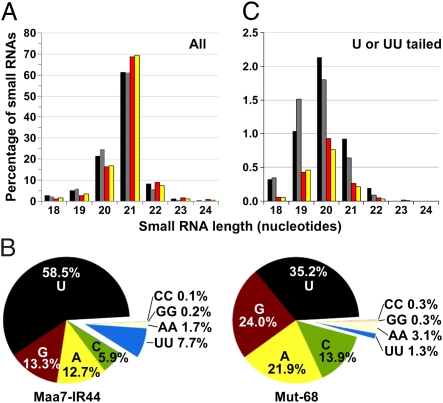
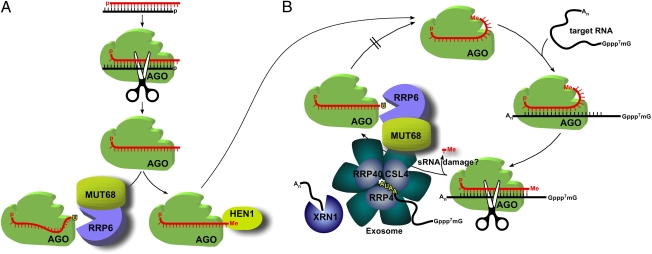
Regulation of gene expression by small RNAs ( approximately 20-30 nucleotides in length) plays an essential role in developmental pathways and defense responses against genomic parasites in eukaryotes. MicroRNAs (miRNAs) and small interfering RNAs (siRNAs) commonly direct the inactivation of cognate sequences through a variety of mechanisms, including RNA degradation, translation inhibition, and transcriptional repression. Recent studies have provided considerable insight into the biogenesis and the mode of action of miRNAs and siRNAs. However, relatively little is known about mechanisms of quality control and small RNA decay in RNA interference (RNAi) pathways. Here we show that deletion of MUT68, encoding a terminal nucleotidyltransferase in the alga Chlamydomonas reinhardtii, results in elevated miRNA and siRNA levels. We found that MUT68 plays a role in the untemplated uridylation of the 3' ends of small RNAs in vivo and stimulates their degradation by the RRP6 exosome subunit in vitro. Moreover, RRP6 depletion also leads to accumulation of small RNAs in vivo. We propose that MUT68 and RRP6 cooperate in the degradation of mature miRNAs and siRNAs, as a quality control mechanism to eliminate dysfunctional or damaged small RNA molecules.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Turnover of mature miRNAs and siRNAs in plants and algae.Adv Exp Med Biol. 2010;700:124-39. Adv Exp Med Biol. 2010. PMID: 21627035 Review.
-
Turnover of Mature miRNAs and siRNAs in Plants and Algae.Adv Exp Med Biol. 2011;700:124-39. doi: 10.1007/978-1-4419-7823-3_11. Adv Exp Med Biol. 2011. PMID: 21755478
-
The Arabidopsis nucleotidyl transferase HESO1 uridylates unmethylated small RNAs to trigger their degradation.Curr Biol. 2012 Apr 24;22(8):689-94. doi: 10.1016/j.cub.2012.02.051. Epub 2012 Mar 29. Curr Biol. 2012. PMID: 22464194 Free PMC article.
-
RNA uridylation and decay in plants.Philos Trans R Soc Lond B Biol Sci. 2018 Nov 5;373(1762):20180163. doi: 10.1098/rstb.2018.0163. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 30397100 Free PMC article. Review.
-
Untemplated oligoadenylation promotes degradation of RISC-cleaved transcripts.Science. 2006 Dec 22;314(5807):1893. doi: 10.1126/science.1135268. Science. 2006. PMID: 17185594
Cited by
-
Systematic Curation of miRBase Annotation Using Integrated Small RNA High-Throughput Sequencing Data for C. elegans and Drosophila.Front Genet. 2011 May 26;2:25. doi: 10.3389/fgene.2011.00025. eCollection 2011. Front Genet. 2011. PMID: 22303321 Free PMC article.
-
Mechanisms of microRNA-mediated gene regulation in unicellular model alga Chlamydomonas reinhardtii.Biotechnol Biofuels. 2018 Sep 8;11:244. doi: 10.1186/s13068-018-1249-y. eCollection 2018. Biotechnol Biofuels. 2018. PMID: 30202439 Free PMC article. Review.
-
The role of 3' end uridylation in RNA metabolism and cellular physiology.Philos Trans R Soc Lond B Biol Sci. 2018 Nov 5;373(1762):20180171. doi: 10.1098/rstb.2018.0171. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 30397107 Free PMC article. Review.
-
New perspectives on the diversification of the RNA interference system: insights from comparative genomics and small RNA sequencing.Wiley Interdiscip Rev RNA. 2014 Mar-Apr;5(2):141-81. doi: 10.1002/wrna.1210. Epub 2013 Dec 5. Wiley Interdiscip Rev RNA. 2014. PMID: 24311560 Free PMC article. Review.
-
Mechanisms of microRNA turnover.Curr Opin Plant Biol. 2015 Oct;27:199-206. doi: 10.1016/j.pbi.2015.07.008. Epub 2015 Sep 2. Curr Opin Plant Biol. 2015. PMID: 26342825 Free PMC article. Review.
References
-
- Carthew RW, Sontheimer EJ. Silence from within: Endogenous siRNAs and miRNAs. Cell. 2009;136:642–655. - PubMed
-
- Baulcombe D. RNA silencing in plants. Nature. 2004;431:356–363. - PubMed
-
- Chapman EJ, Carrington JC. Specialization and evolution of endogenous small RNA pathways. Nat Rev Genet. 2007;8:884–896. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
