

Loss of myeloid cell-derived vascular endothelial growth factor accelerates fibrosis
- PMID: 20142499
- PMCID: PMC2840078
- DOI: 10.1073/pnas.0912766107
Loss of myeloid cell-derived vascular endothelial growth factor accelerates fibrosis
Abstract
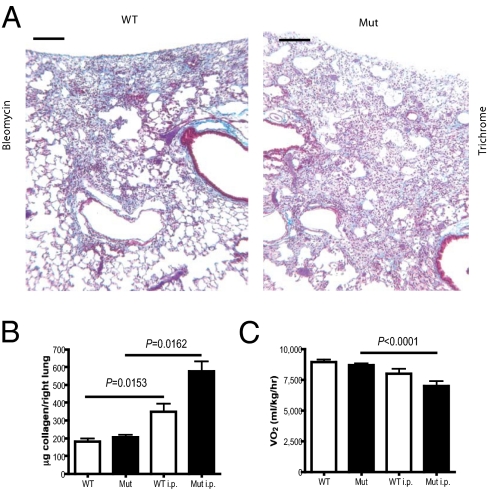
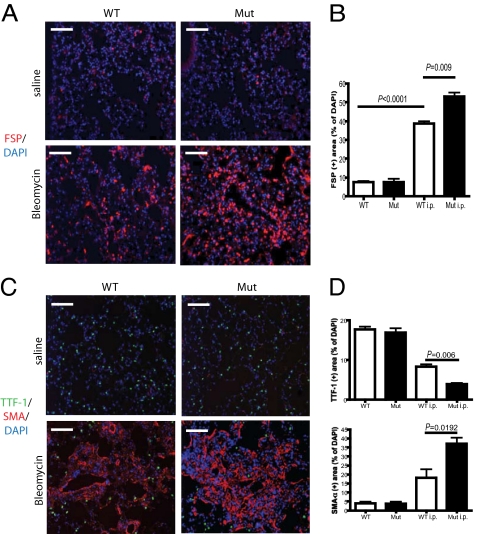
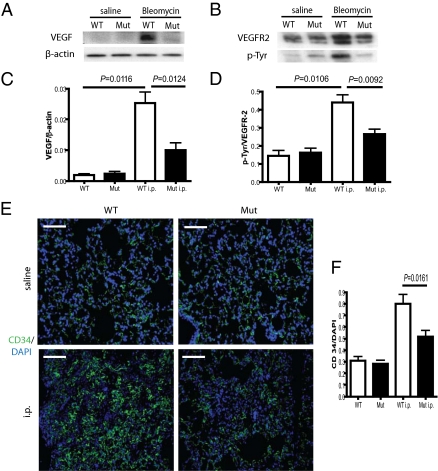
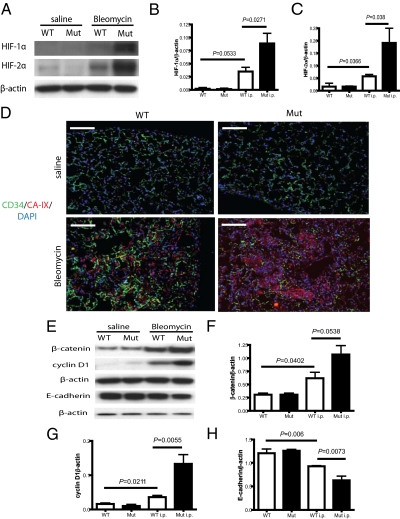
Tissue injury initiates a complex series of events that act to restore structure and physiological homeostasis. Infiltration of inflammatory cells and vascular remodeling are both keystones of this process. However, the role of inflammation and angiogenesis in general and, more specifically, the significance of inflammatory cell-derived VEGF in this context are unclear. To determine the role of inflammatory cell-derived VEGF in a clinically relevant and chronically inflamed injury, pulmonary fibrosis, we deleted the VEGF-A gene in myeloid cells. In a model of pulmonary fibrosis in mice, deletion of VEGF in myeloid cells resulted in significantly reduced formation of blood vessels; however, it causes aggravated fibrotic tissue damage. This was accompanied by a pronounced decrease in epithelial cell survival and a striking increase in myofibroblast invasion. The drastic increase in fibrosis following loss of myeloid VEGF in the damaged lungs was also marked by increased levels of hypoxia-inducible factor (HIF) expression and Wnt/beta-catenin signaling. This demonstrates that the process of angiogenesis, driven by myeloid cell-derived VEGF, is essential for the prevention of fibrotic damage.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Deletion of vascular endothelial growth factor in myeloid cells accelerates tumorigenesis.Nature. 2008 Dec 11;456(7223):814-8. doi: 10.1038/nature07445. Epub 2008 Nov 9. Nature. 2008. PMID: 18997773 Free PMC article.
-
[Relationship between bleomycin-induced pulmonary fibrosis and vascular endothelial cell injury].Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi. 2004 Oct;22(5):354-7. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi. 2004. PMID: 15555311 Chinese.
-
VEGFR-2 antagonist SU5416 attenuates bleomycin-induced pulmonary fibrosis in mice.Int Immunopharmacol. 2009 Jan;9(1):70-9. doi: 10.1016/j.intimp.2008.10.002. Epub 2008 Oct 29. Int Immunopharmacol. 2009. PMID: 18976720
-
Chemokine signaling axis between endothelial and myeloid cells regulates development of pulmonary hypertension associated with pulmonary fibrosis and hypoxia.Am J Physiol Lung Cell Mol Physiol. 2019 Oct 1;317(4):L434-L444. doi: 10.1152/ajplung.00156.2019. Epub 2019 Jul 31. Am J Physiol Lung Cell Mol Physiol. 2019. PMID: 31364370 Free PMC article.
-
Fell-Muir Lecture: Regulatory mechanisms of skeletal and connective tissue development and homeostasis - lessons from studies of human disorders.Int J Exp Pathol. 2016 Aug;97(4):296-302. doi: 10.1111/iep.12198. Epub 2016 Sep 1. Int J Exp Pathol. 2016. PMID: 27581728 Free PMC article. Review.
Cited by
-
Therapeutic efficacy of thrombin-preconditioned mesenchymal stromal cell-derived extracellular vesicles on Escherichia coli-induced acute lung injury in mice.Respir Res. 2024 Aug 7;25(1):303. doi: 10.1186/s12931-024-02908-w. Respir Res. 2024. PMID: 39112999 Free PMC article.
-
VEGF (Vascular Endothelial Growth Factor) and Fibrotic Lung Disease.Int J Mol Sci. 2018 Apr 24;19(5):1269. doi: 10.3390/ijms19051269. Int J Mol Sci. 2018. PMID: 29695053 Free PMC article. Review.
-
RNA-sequencing profiling analysis of pericyte-derived extracellular vesicle-mimetic nanovesicles-regulated genes in primary cultured fibroblasts from normal and Peyronie's disease penile tunica albuginea.BMC Urol. 2021 Aug 6;21(1):103. doi: 10.1186/s12894-021-00872-x. BMC Urol. 2021. PMID: 34362357 Free PMC article.
-
Myeloid-Epithelial-Reproductive Receptor Tyrosine Kinase and Milk Fat Globule Epidermal Growth Factor 8 Coordinately Improve Remodeling After Myocardial Infarction via Local Delivery of Vascular Endothelial Growth Factor.Circulation. 2016 Mar 1;133(9):826-39. doi: 10.1161/CIRCULATIONAHA.115.020857. Epub 2016 Jan 27. Circulation. 2016. PMID: 26819373 Free PMC article.
-
[An update of understanding of the hepatic vascular system and new research strategies].Nan Fang Yi Ke Da Xue Xue Bao. 2022 Dec 20;42(12):1907-1911. doi: 10.12122/j.issn.1673-4254.2022.12.22. Nan Fang Yi Ke Da Xue Xue Bao. 2022. PMID: 36651262 Free PMC article. Review. Chinese.
References
-
- Selman M, King TE, Pardo A, American Thoracic Society. European Respiratory Society. American College of Chest Physicians Idiopathic pulmonary fibrosis: Prevailing and evolving hypotheses about its pathogenesis and implications for therapy. Ann Intern Med. 2001;134:136–151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
