

Peroxynitrite-mediated oxidative modifications of complex II: relevance in myocardial infarction
- PMID: 20143804
- PMCID: PMC3754874
- DOI: 10.1021/bi9018237
Peroxynitrite-mediated oxidative modifications of complex II: relevance in myocardial infarction
Abstract
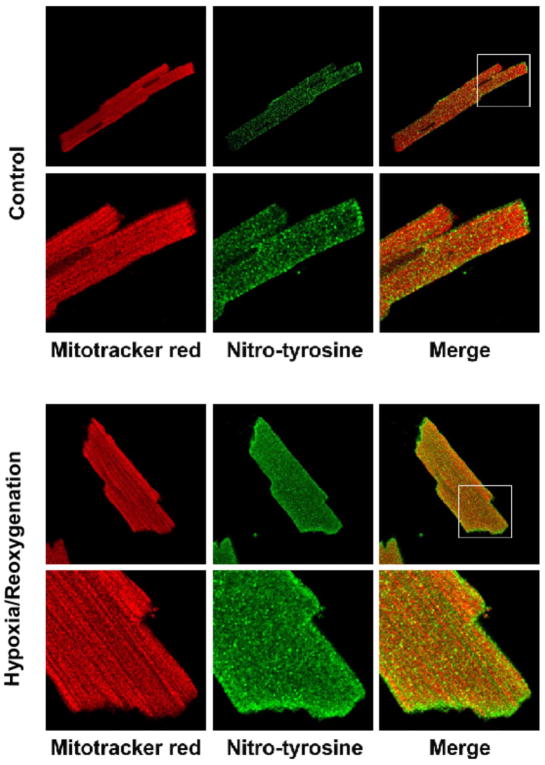
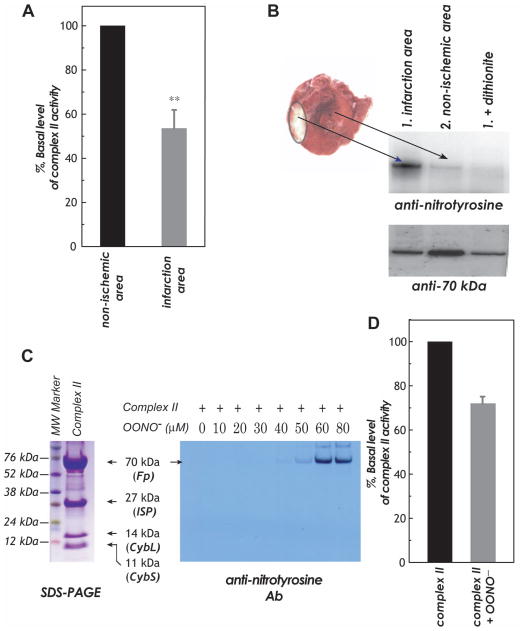
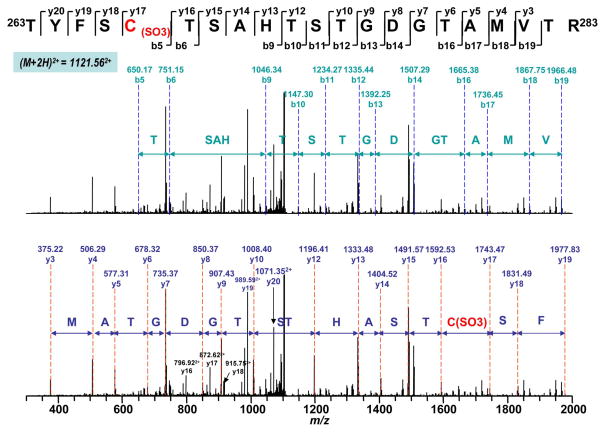
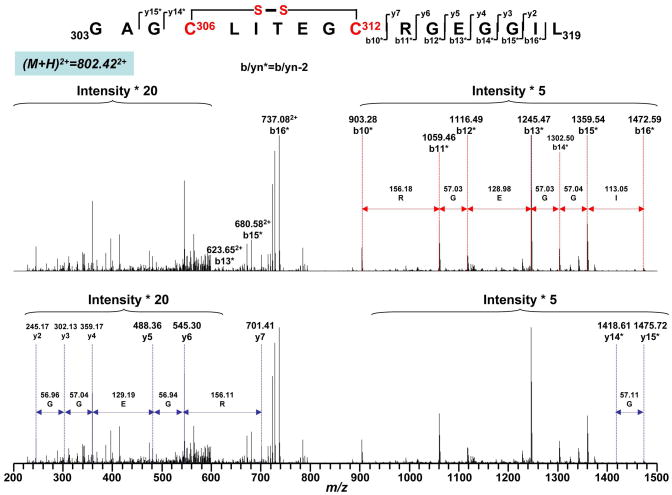
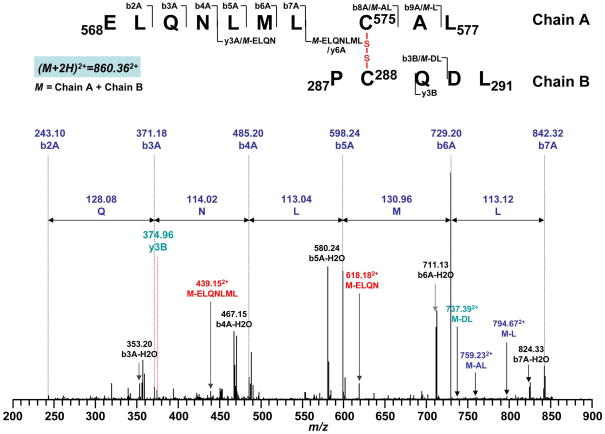
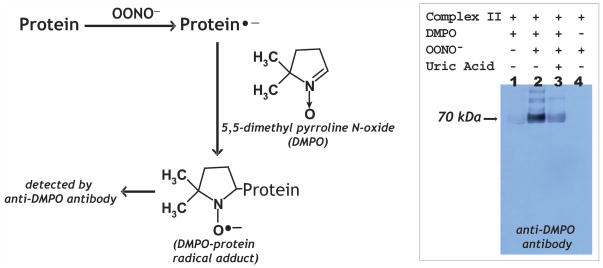
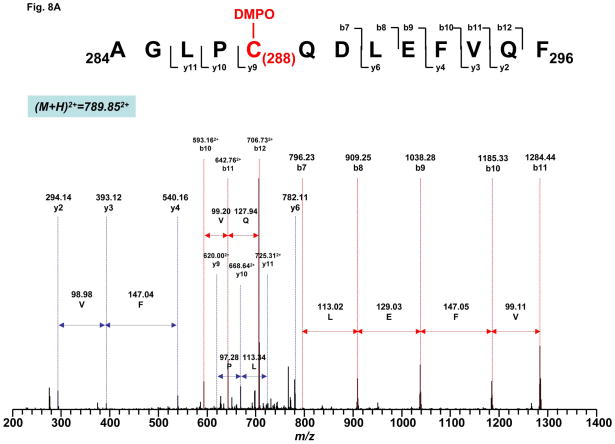
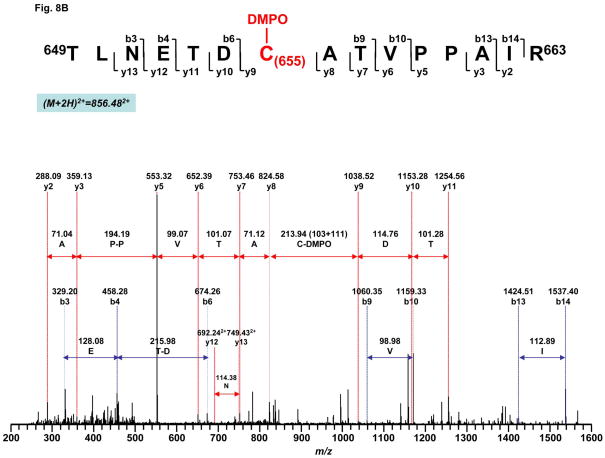
Increased O(2)(*-) and NO production is a key mechanism of mitochondrial dysfunction in myocardial ischemia/reperfusion injury. In complex II, oxidative impairment and enhanced tyrosine nitration of the 70 kDa FAD-binding protein occur in the post-ischemic myocardium and are thought to be mediated by peroxynitrite (OONO(-)) in vivo [Chen, Y.-R., et al. (2008) J. Biol. Chem. 283, 27991-28003]. To gain deeper insights into the redox protein thiols involved in OONO(-)-mediated oxidative post-translational modifications relevant in myocardial infarction, we subjected isolated myocardial complex II to in vitro protein nitration with OONO(-). This resulted in site-specific nitration at the 70 kDa polypeptide and impairment of complex II-derived electron transfer activity. Under reducing conditions, the gel band of the 70 kDa polypeptide was subjected to in-gel trypsin/chymotrypsin digestion and then LC-MS/MS analysis. Nitration of Y(56) and Y(142) was previously reported. Further analysis revealed that C(267), C(476), and C(537) are involved in OONO(-)-mediated S-sulfonation. To identify the disulfide formation mediated by OONO(-), nitrated complex II was alkylated with iodoacetamide. In-gel proteolytic digestion and LC-MS/MS analysis were conducted under nonreducing conditions. The MS/MS data were examined with MassMatrix, indicating that three cysteine pairs, C(306)-C(312), C(439)-C(444), and C(288)-C(575), were involved in OONO(-)-mediated disulfide formation. Immuno-spin trapping with an anti-DMPO antibody and subsequent MS was used to define oxidative modification with protein radical formation. An OONO(-)-dependent DMPO adduct was detected, and further LC-MS/MS analysis indicated C(288) and C(655) were involved in DMPO binding. These results offered a complete profile of OONO(-)-mediated oxidative modifications that may be relevant in the disease model of myocardial infarction.
Figures

References
-
- Lemos RS, Fernandes AS, Pereira MM, Gomes CM, Teixeira M. Quinol:fumarate oxidoreductases and succinate:quinone oxidoreductases: phylogenetic relationships, metal centres and membrane attachment. Biochim Biophys Acta. 2002;1553:158–170. - PubMed
-
- Chen YR, Chen CL, Pfeiffer DR, Zweier JL. Mitochondrial Complex II in the Post-ischemic Heart: OXIDATIVE INJURY AND THE ROLE OF PROTEIN S-GLUTATHIONYLATION. J Biol Chem. 2007;282:32640–32654. - PubMed
-
- Zhao X, Chen YR, He G, Zhang A, Druhan LJ, Strauch AR, Zweier JL. Endothelial nitric oxide synthase (NOS3) knockout decreases NOS2 induction, limiting hyperoxygenation and conferring protection in the postischemic heart. Am J Physiol Heart Circ Physiol. 2007;292:H1541–1550. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
