

Vimentin is a functional partner of hormone sensitive lipase and facilitates lipolysis
- PMID: 20143880
- PMCID: PMC2849902
- DOI: 10.1021/pr900909t
Vimentin is a functional partner of hormone sensitive lipase and facilitates lipolysis
Abstract
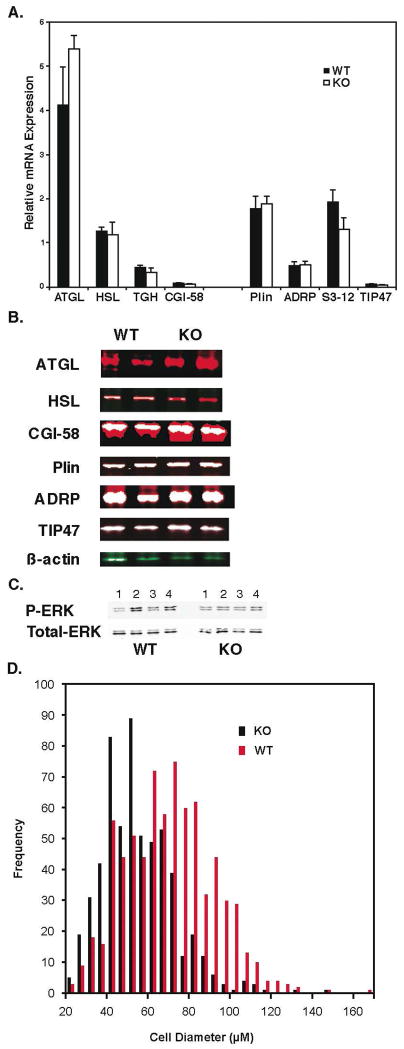
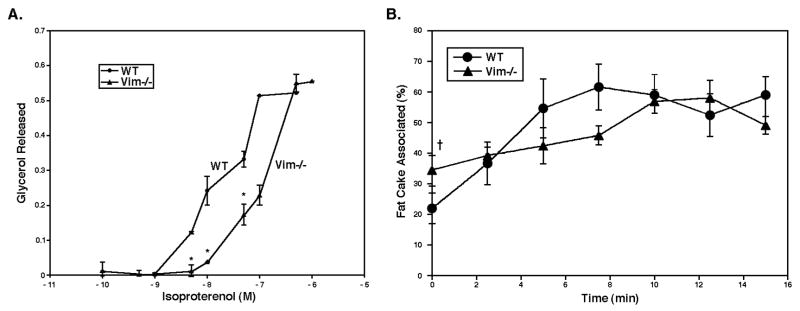
Lipolysis involves a number of components including signaling pathways, droplet-associated proteins, and lipases such as hormone-sensitive lipase (HSL). We used surface enhanced laser desorption/ionization time-of-flight mass spectroscopy to identify cellular proteins that might interact with HSL and potentially influence lipolysis. Using recombinant HSL as bait on protein chips, clusters of proteins of 14.7-18.9, 25.8-26.8, 36.1, 44.3-49.1, and 53.7 kDa were identified that interact with HSL, particularly when lysates were examined from beta-agonist treated mouse adipocytes. The ability to detect these interacting proteins was markedly diminished when the adipocytes were treated with insulin. A very similar pattern of proteins was identified when anti-HSL IgG was used as the bait. Following immunocapture, the identification of the prominent 53.7 kDa protein was carried out by tryptic digestion and MS analysis and determined to be vimentin. The interaction of HSL with vimentin, and its hormonal dependence, was confirmed by coimmunoprecipitation. beta-Agonist stimulated lipolysis and the rate of HSL translocation were impaired in vimentin null adipocytes, even though normal amounts of lipases and droplet-associated proteins are expressed. The current studies provide evidence that vimentin participates in lipolysis through direct, hormonally regulated interactions with HSL.
Figures

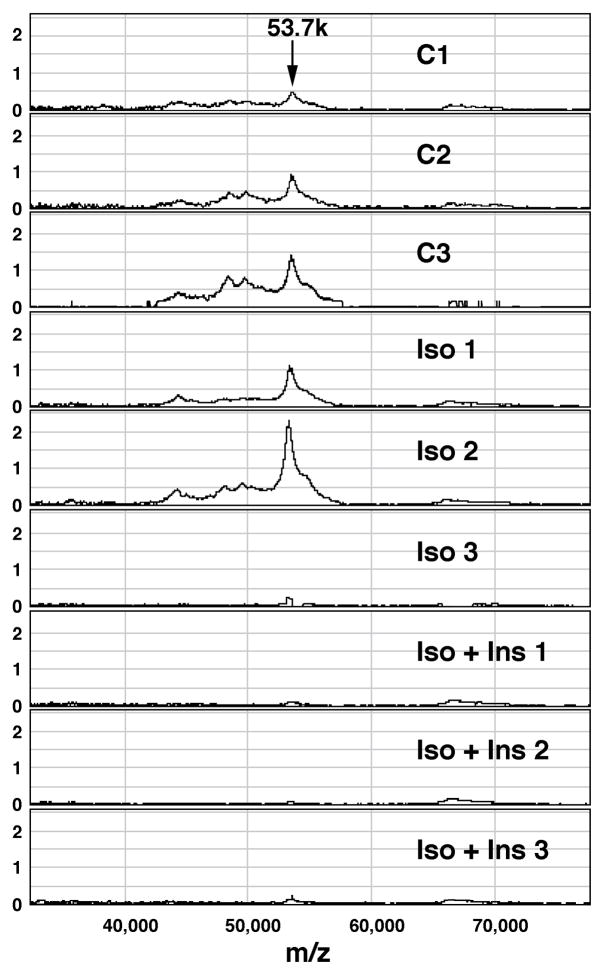
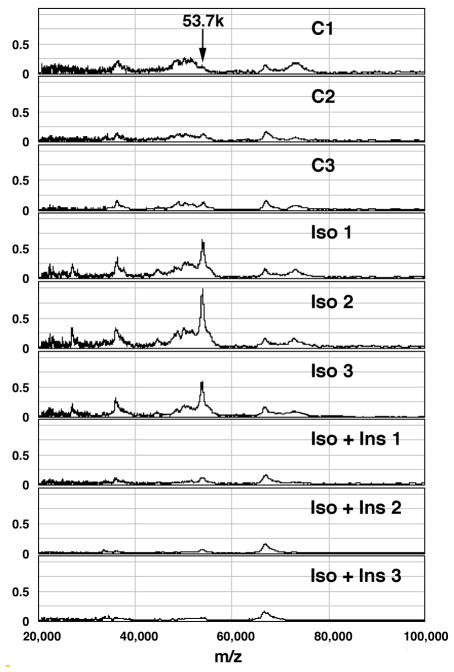
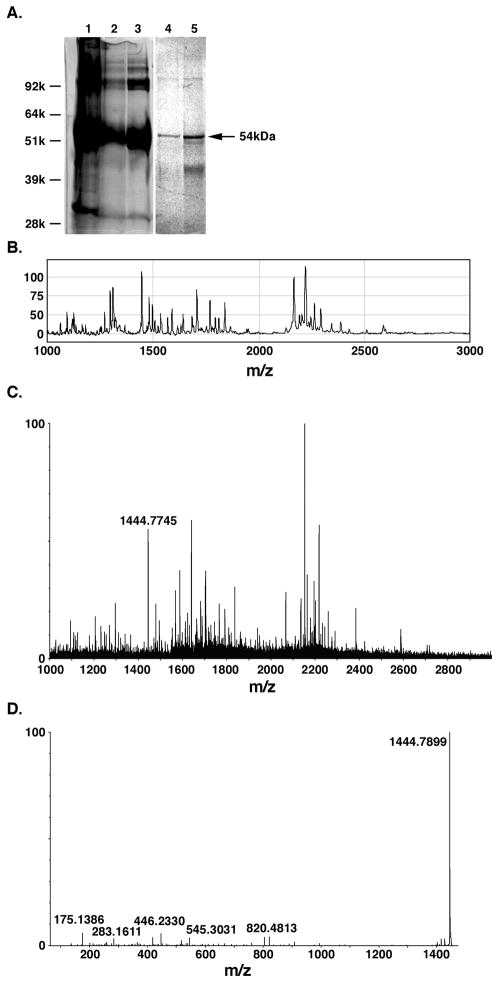
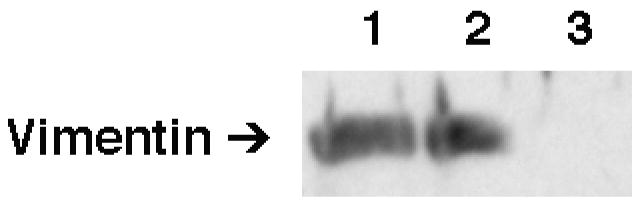
Similar articles
-
Perilipin promotes hormone-sensitive lipase-mediated adipocyte lipolysis via phosphorylation-dependent and -independent mechanisms.J Biol Chem. 2006 Jun 9;281(23):15837-44. doi: 10.1074/jbc.M601097200. Epub 2006 Apr 4. J Biol Chem. 2006. PMID: 16595669
-
Lipolysis in the absence of hormone-sensitive lipase: evidence for a common mechanism regulating distinct lipases.Diabetes. 2002 Dec;51(12):3368-75. doi: 10.2337/diabetes.51.12.3368. Diabetes. 2002. PMID: 12453888
-
Functional interaction of hormone-sensitive lipase and perilipin in lipolysis.J Lipid Res. 2009 Nov;50(11):2306-13. doi: 10.1194/jlr.M900176-JLR200. Epub 2009 Jun 10. J Lipid Res. 2009. PMID: 19515989 Free PMC article.
-
Molecular mechanisms regulating hormone-sensitive lipase and lipolysis.Biochem Soc Trans. 2003 Dec;31(Pt 6):1120-4. doi: 10.1042/bst0311120. Biochem Soc Trans. 2003. PMID: 14641008 Review.
-
An Overview of Hormone-Sensitive Lipase (HSL).ScientificWorldJournal. 2022 Dec 8;2022:1964684. doi: 10.1155/2022/1964684. eCollection 2022. ScientificWorldJournal. 2022. PMID: 36530555 Free PMC article. Review.
Cited by
-
Plasma Cytokeratin-18 Fragment Level Reflects the Metabolic Phenotype in Obesity.Biomolecules. 2023 Apr 14;13(4):675. doi: 10.3390/biom13040675. Biomolecules. 2023. PMID: 37189422 Free PMC article.
-
Altered Protein Composition of Subcutaneous Adipose Tissue in Chronic Kidney Disease.Kidney Int Rep. 2017 Jul 29;2(6):1208-1218. doi: 10.1016/j.ekir.2017.07.007. eCollection 2017 Nov. Kidney Int Rep. 2017. PMID: 29270529 Free PMC article.
-
cAMP-stimulated phosphorylation of diaphanous 1 regulates protein stability and interaction with binding partners in adrenocortical cells.Mol Biol Cell. 2013 Mar;24(6):848-57. doi: 10.1091/mbc.E12-08-0597. Epub 2013 Jan 16. Mol Biol Cell. 2013. PMID: 23325789 Free PMC article.
-
Proteome Profile and Quantitative Proteomic Analysis of Buffalo (Bubalusbubalis) Follicular Fluid during Follicle Development.Int J Mol Sci. 2016 Apr 29;17(5):618. doi: 10.3390/ijms17050618. Int J Mol Sci. 2016. PMID: 27136540 Free PMC article.
-
Melanoma cells induce dedifferentiation and metabolic changes in adipocytes present in the tumor niche.Cell Mol Biol Lett. 2023 Jul 22;28(1):58. doi: 10.1186/s11658-023-00476-3. Cell Mol Biol Lett. 2023. PMID: 37481560 Free PMC article.
References
-
- Greenberg AS, Shen W-J, Muliro K, Patel S, Souza SC, Roth RA, Kraemer FB. Stimulation of lipolysis and hormone-sensitive lipase via the extracellular signal-regulated kinase pathway. J Biol Chem. 2001;276(48):45456–45461. - PubMed
-
- Robidoux J, Kumar N, Daniel KW, Moukdar F, Cyr M, Medvedev AV, Collins S. Maximal beta3-adrenergic regulation of lipolysis involves Src and epidermal growth factor receptor-dependent ERK1/2 activation. J Biol Chem. 2006;281(49):37794–802. - PubMed
-
- Kraemer FB, Shen W-J. Hormone-sensitive lipase: control of intracellular tri-(di-) acylglycerol and cholesteryl ester hydrolysis. J Lipid Res. 2002;43(10):1585–1594. - PubMed
-
- Greenberg AS, Egan JJ, Wek SA, Garty NB, Blanchette-Mackie EJ, Londos C. Perilipin, a major hormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storage droplets. J Biol Chem. 1991;266(17):11341–11346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
