

Binding of NF-kappaB p65 subunit to the promoter elements is involved in LPS-induced transactivation of miRNA genes in human biliary epithelial cells
- PMID: 20144951
- PMCID: PMC2879527
- DOI: 10.1093/nar/gkq056
Binding of NF-kappaB p65 subunit to the promoter elements is involved in LPS-induced transactivation of miRNA genes in human biliary epithelial cells
Abstract
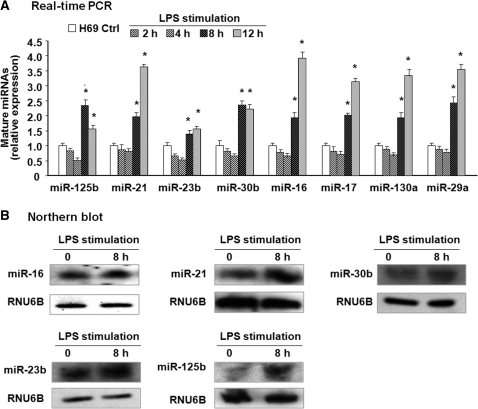
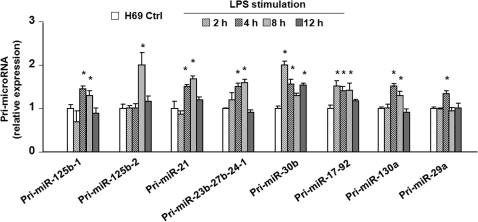
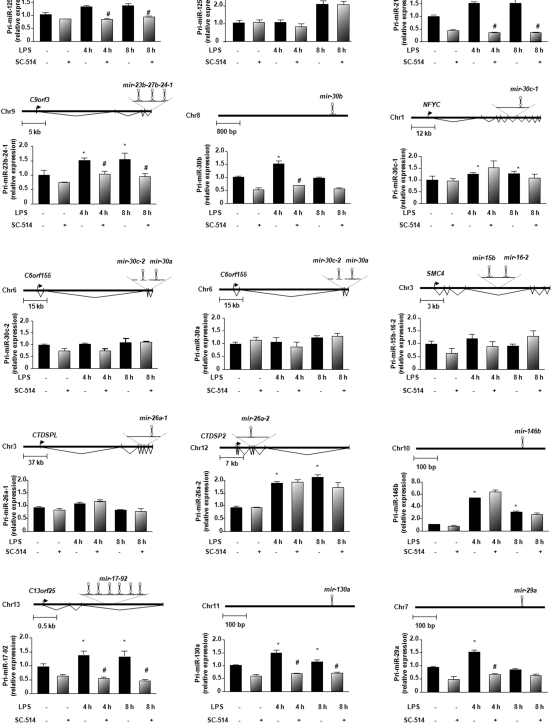
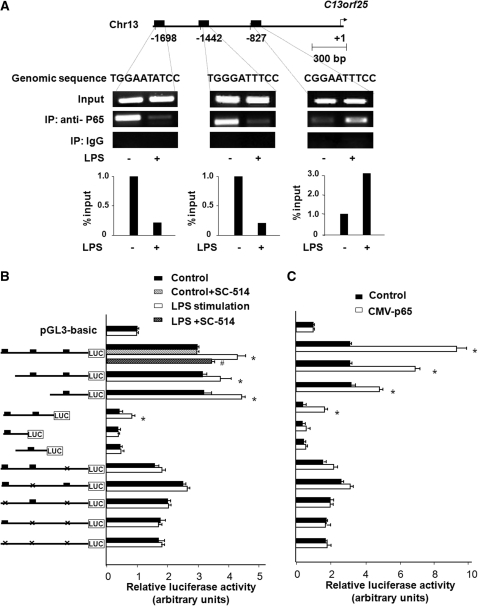
The majority of human miRNA genes is transcribed by polymerase II and can be classified as class II genes similar to protein-coding genes. Whereas current research on miRNAs has focused on the physiological and pathological functions, the molecular mechanisms underlying their transcriptional regulation are largely unknown. We recently reported that lipopolysaccharide (LPS) alters mature miRNA expression profile in human biliary epithelial cells. In this study, we tested the role of transcription factor NF-kappaB in LPS-induced transcription of select miRNA genes. Of the majority of LPS-up-regulated mature miRNAs in cultured human biliary epithelial cells, potential NF-kappaB binding sites were identified in the putative promoter elements of their corresponding genes. Inhibition of NF-kappaB activation by SC-514, an IKK2 inhibitor, blocked LPS-induced up-regulation of a subset of pri-miRNAs, including pri-miR-17-92, pri-miR-125b-1, pri-miR-21, pri-miR-23b-27b-24-1, pri-miR-30b, pri-miR-130a and pri-miR-29a. Moreover, direct binding of NF-kappaB p65 subunit to the promoter elements of mir-17-92, mir-125b-1, mir-21, mir-23b-27b-24-1, mir-30b and mir-130a genes was identified by chromatin immunoprecipitation analysis and confirmed by the luciferase reporter assay. Thus, a subset of miRNA genes is regulated in human biliary epithelial cells through NF-kappaB activation induced by LPS, suggesting a role of the NF-kappaB pathway in the transcriptional regulation of miRNA genes.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
