

Targeting of the cytosolic poly(A) binding protein PABPC1 to mitochondria causes mitochondrial translation inhibition
- PMID: 20144953
- PMCID: PMC2887948
- DOI: 10.1093/nar/gkq068
Targeting of the cytosolic poly(A) binding protein PABPC1 to mitochondria causes mitochondrial translation inhibition
Abstract
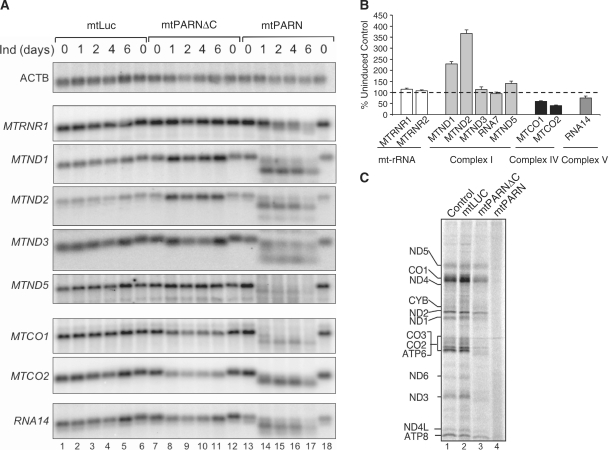
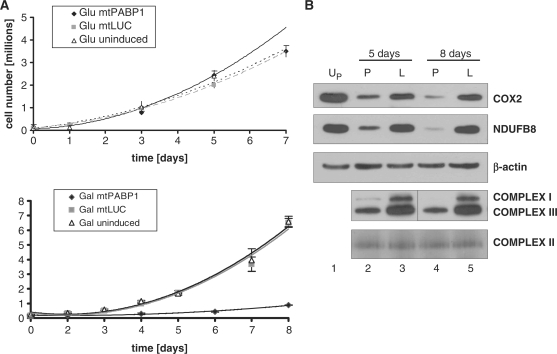
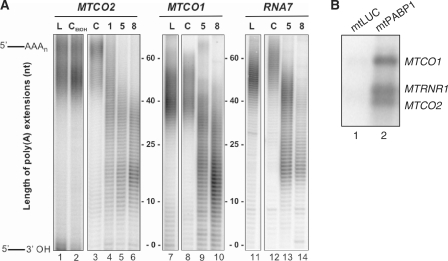
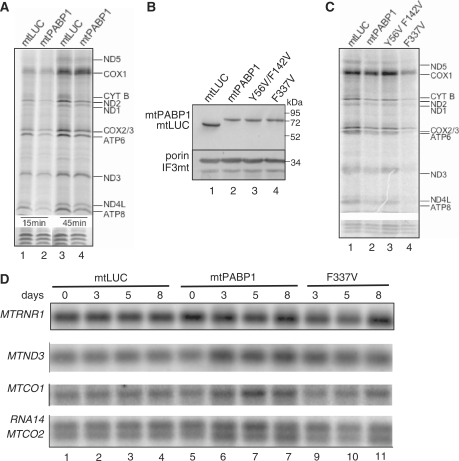
Mammalian mitochondria contain their own genome that is almost fully transcribed from both strands, generating polycistronic RNA units that are processed and matured. The mitochondrial mRNA is modified by oligo- or polyadenylation at the 3' termini, but the exact function of this post-transcriptional addition is unclear. Current debate focuses on the role of polyadenylation in transcript stability. An equally likely function that has received little attention is that, as in the cytosol of eukaryotes, polyadenylation facilitates translation in the mitochondrion. To address this issue, we have targeted cytosolic proteins to the mitochondrion, a poly(A) specific 3' exoribonuclease, mtPARN, and a poly(A)binding protein, mtPABP1. Removal of the 3' adenylyl extensions had a variable effect on mt-mRNA steady-state levels, increasing (MTND1, 2, 5) or decreasing (MTCO1, 2, RNA14) certain species with minimal effect on others (RNA7, MTND3). Translation was markedly affected, but interpretation of this was complicated by the concomitant 3' truncation of the open reading frame in most cases. Coating of the poly(A) tail by mtPABP1, however, did not lead to transcript decay but caused a marked inhibition of mitochondrial translation. These data are consistent with endogenous RNA-binding factor(s) interacting with the poly(A) to optimize mitochondrial protein synthesis.
Figures
Similar articles
-
Investigation of a pathogenic mtDNA microdeletion reveals a translation-dependent deadenylation decay pathway in human mitochondria.Hum Mol Genet. 2003 Sep 15;12(18):2341-8. doi: 10.1093/hmg/ddg238. Epub 2003 Jul 22. Hum Mol Genet. 2003. PMID: 12915481
-
PDE12 removes mitochondrial RNA poly(A) tails and controls translation in human mitochondria.Nucleic Acids Res. 2011 Sep 1;39(17):7750-63. doi: 10.1093/nar/gkr470. Epub 2011 Jun 11. Nucleic Acids Res. 2011. PMID: 21666256 Free PMC article.
-
Maturation of selected human mitochondrial tRNAs requires deadenylation.Elife. 2017 Jul 26;6:e27596. doi: 10.7554/eLife.27596. Elife. 2017. PMID: 28745585 Free PMC article.
-
PARN-like Proteins Regulate Gene Expression in Land Plant Mitochondria by Modulating mRNA Polyadenylation.Int J Mol Sci. 2021 Oct 5;22(19):10776. doi: 10.3390/ijms221910776. Int J Mol Sci. 2021. PMID: 34639116 Free PMC article. Review.
-
RNA degradation in human mitochondria: the journey is not finished.Hum Mol Genet. 2024 May 22;33(R1):R26-R33. doi: 10.1093/hmg/ddae043. Hum Mol Genet. 2024. PMID: 38779774 Free PMC article. Review.
Cited by
-
Messenger RNA delivery to mitoribosomes - hints from a bacterial toxin.FEBS J. 2021 Jan;288(2):437-451. doi: 10.1111/febs.15342. Epub 2020 May 11. FEBS J. 2021. PMID: 32329962 Free PMC article.
-
Alternative translation initiation augments the human mitochondrial proteome.Nucleic Acids Res. 2013 Feb 1;41(4):2354-69. doi: 10.1093/nar/gks1347. Epub 2012 Dec 28. Nucleic Acids Res. 2013. PMID: 23275553 Free PMC article.
-
Mitochondrial transcription and translation: overview.Essays Biochem. 2018 Jul 20;62(3):309-320. doi: 10.1042/EBC20170102. Print 2018 Jul 20. Essays Biochem. 2018. PMID: 30030363 Free PMC article. Review.
-
SUV3 helicase is required for correct processing of mitochondrial transcripts.Nucleic Acids Res. 2015 Sep 3;43(15):7398-413. doi: 10.1093/nar/gkv692. Epub 2015 Jul 7. Nucleic Acids Res. 2015. PMID: 26152302 Free PMC article.
-
Defective mitochondrial mRNA maturation is associated with spastic ataxia.Am J Hum Genet. 2010 Nov 12;87(5):655-60. doi: 10.1016/j.ajhg.2010.09.013. Epub 2010 Oct 21. Am J Hum Genet. 2010. PMID: 20970105 Free PMC article.
References
-
- Anderson S, Bankier AT, Barrell BG, De Bruijn M.HL, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, et al. Sequence and organization of the human mitochondrial genome. Nature. 1981;290:457–465. - PubMed
-
- Ojala D, Montoya J, Attardi G. tRNA punctuation model of RNA processing in human mitochondria. Nature. 1981;290:470–474. - PubMed
-
- Gagliardi D, Stepien PP, Temperley RJ, Lightowlers RN, Chrzanowska-Lightowlers ZM. Messenger RNA stability in mitochondria: different means to an end. Trends Genet. 2004;20:260–267. - PubMed
-
- Schuster G, Stern D. RNA polyadenylation and decay in mitochondria and chloroplasts. Prog. Mol. Biol. Transl. Sci. 2009;85:393–422. - PubMed
