

Pharmacological prion protein silencing accelerates central nervous system autoimmune disease via T cell receptor signalling
- PMID: 20145049
- PMCID: PMC2822628
- DOI: 10.1093/brain/awp298
Pharmacological prion protein silencing accelerates central nervous system autoimmune disease via T cell receptor signalling
Abstract
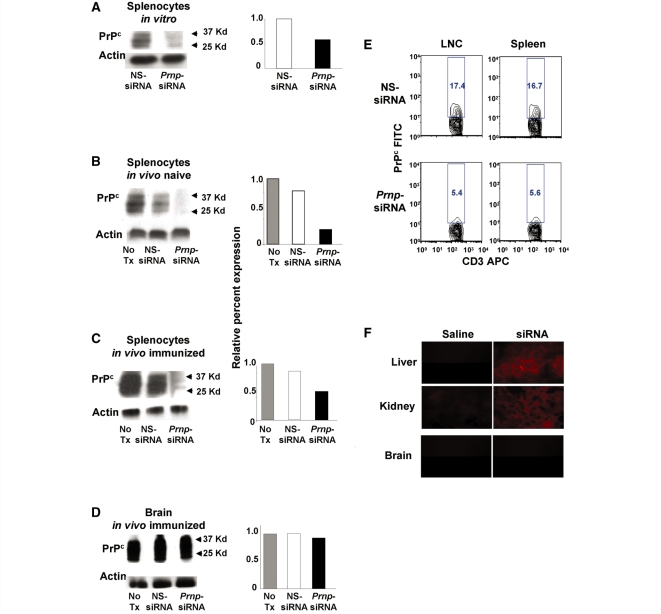
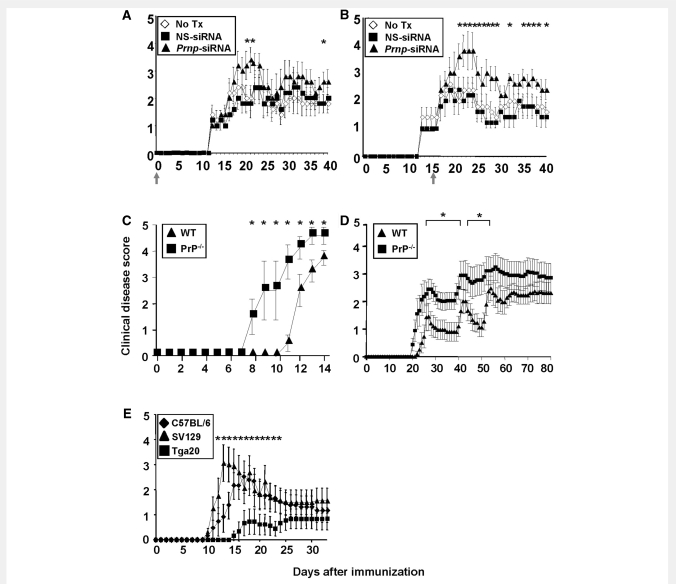
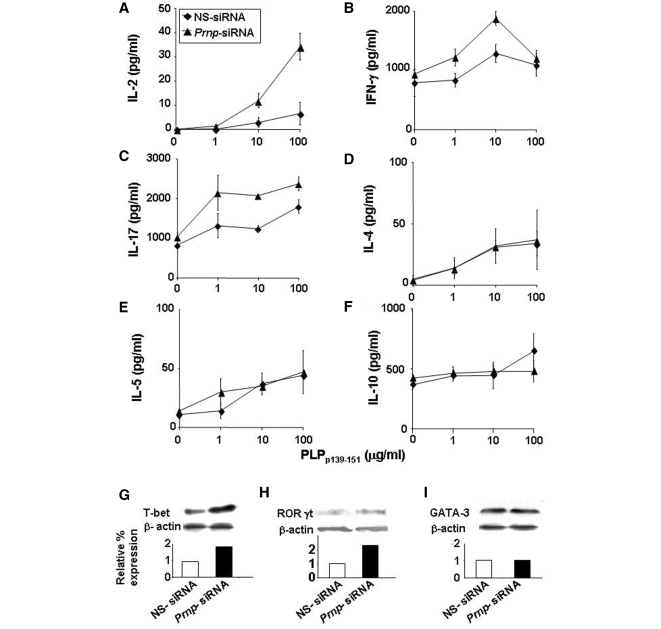
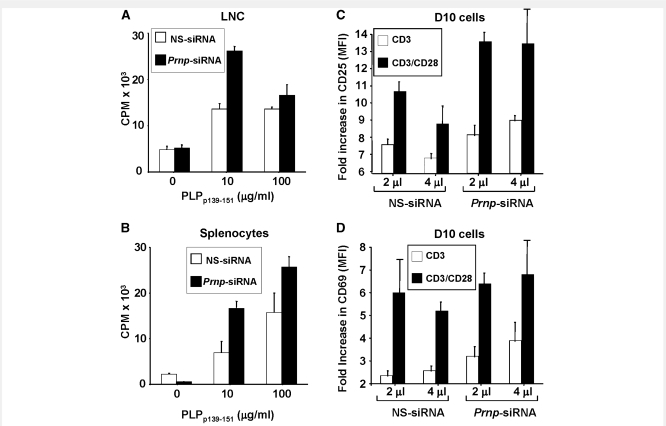
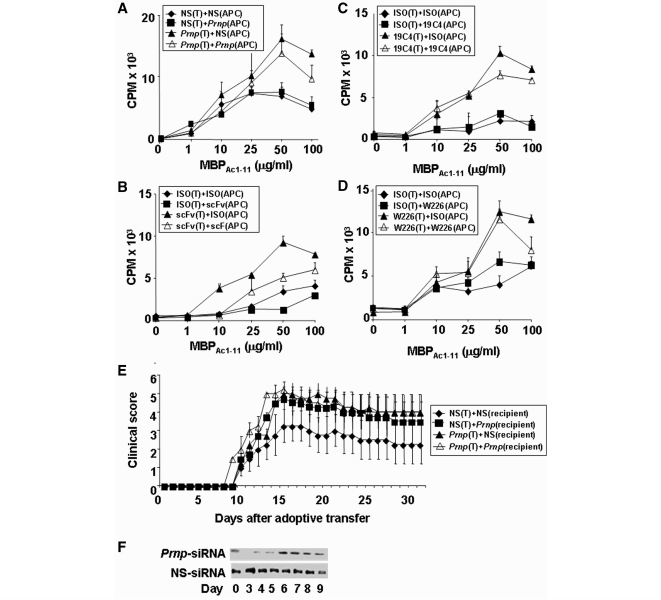
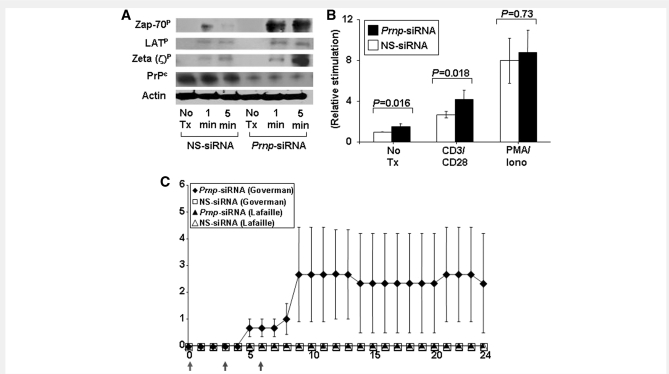
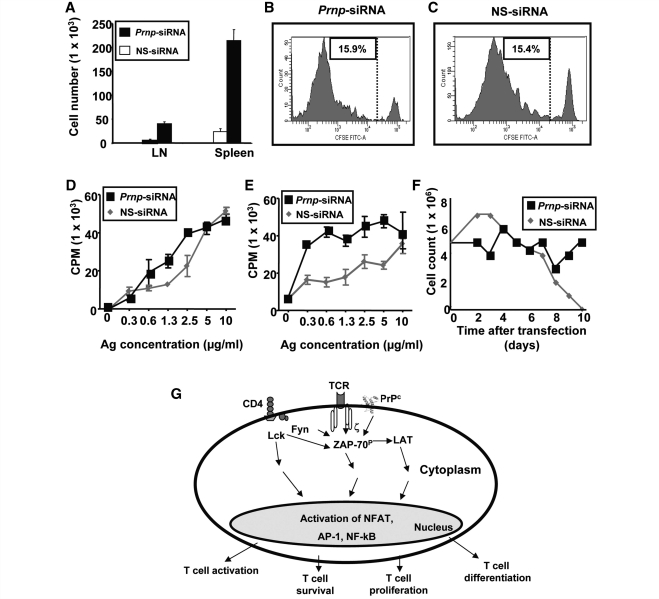
The primary biological function of the endogenous cellular prion protein has remained unclear. We investigated its biological function in the generation of cellular immune responses using cellular prion protein gene-specific small interfering ribonucleic acid in vivo and in vitro. Our results were confirmed by blocking cellular prion protein with monovalent antibodies and by using cellular prion protein-deficient and -transgenic mice. In vivo prion protein gene-small interfering ribonucleic acid treatment effects were of limited duration, restricted to secondary lymphoid organs and resulted in a 70% reduction of cellular prion protein expression in leukocytes. Disruption of cellular prion protein signalling augmented antigen-specific activation and proliferation, and enhanced T cell receptor signalling, resulting in zeta-chain-associated protein-70 phosphorylation and nuclear factor of activated T cells/activator protein 1 transcriptional activity. In vivo prion protein gene-small interfering ribonucleic acid treatment promoted T cell differentiation towards pro-inflammatory phenotypes and increased survival of antigen-specific T cells. Cellular prion protein silencing with small interfering ribonucleic acid also resulted in the worsening of actively induced and adoptively transferred experimental autoimmune encephalomyelitis. Finally, treatment of myelin basic protein(1-11) T cell receptor transgenic mice with prion protein gene-small interfering ribonucleic acid resulted in spontaneous experimental autoimmune encephalomyelitis. Thus, central nervous system autoimmune disease was modulated at all stages of disease: the generation of the T cell effector response, the elicitation of T effector function and the perpetuation of cellular immune responses. Our findings indicate that cellular prion protein regulates T cell receptor-mediated T cell activation, differentiation and survival. Defects in autoimmunity are restricted to the immune system and not the central nervous system. Our data identify cellular prion protein as a regulator of cellular immunological homoeostasis and suggest cellular prion protein as a novel potential target for therapeutic immunomodulation.
Figures
References
-
- Antoine N, Cesbron JY, Coumans B, Jolois O, Zorzi W, Heinen E. Differential expression of cellular prion protein on human blood and tonsil lymphocytes. Haematologica. 2000;85:475–80. - PubMed
-
- Bueler H, Fischer M, Lang Y, Bluethmann H, Lipp HP, DeArmond SJ, et al. Normal development and behaviour of mice lacking the neuronal cell-surface PrP protein. Nature. 1992;356:577–82. - PubMed
-
- Burthem J, Urban B, Pain A, Roberts DJ. The normal cellular prion protein is strongly expressed by myeloid dendritic cells. Blood. 2001;98:3733–8. - PubMed
-
- Cashman NR, Loertscher R, Nalbantoglu J, Shaw I, Kascsak RJ, Bolton DC, et al. Cellular isoform of the scrapie agent protein participates in lymphocyte activation. Cell. 1990;61:185–92. - PubMed
-
- Chan AC, Iwashima M, Turck CW, Weiss A. ZAP-70: a 70 kd protein-tyrosine kinase that associates with the TCR zeta chain. Cell. 1992;71:649–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
