

Very-long-chain fatty acids are involved in polar auxin transport and developmental patterning in Arabidopsis
- PMID: 20145257
- PMCID: PMC2845409
- DOI: 10.1105/tpc.109.071209
Very-long-chain fatty acids are involved in polar auxin transport and developmental patterning in Arabidopsis
Abstract
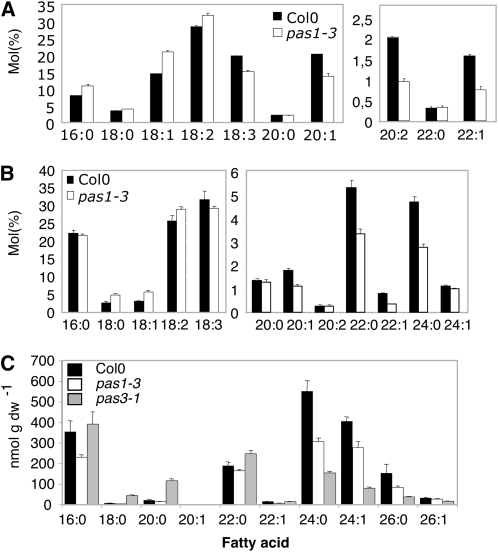
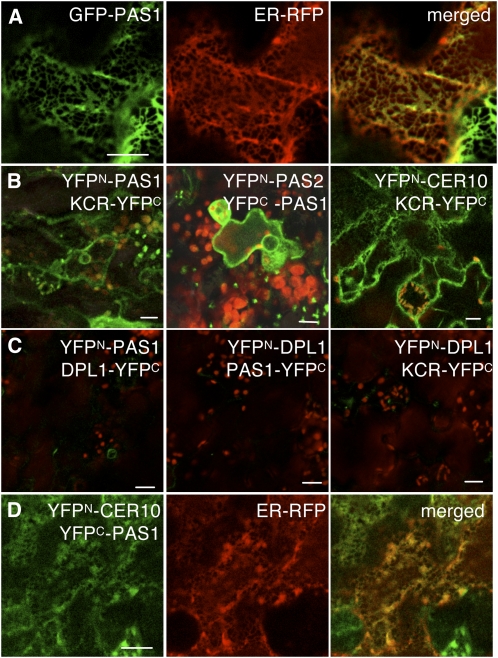
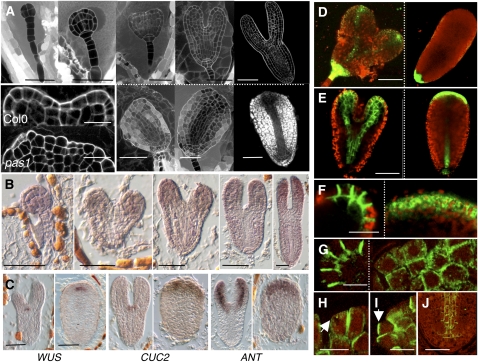
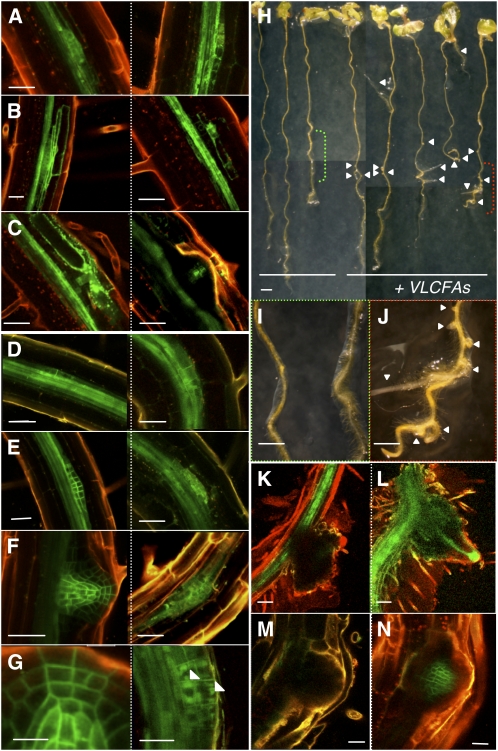
Very-long-chain fatty acids (VLCFAs) are essential for many aspects of plant development and necessary for the synthesis of seed storage triacylglycerols, epicuticular waxes, and sphingolipids. Identification of the acetyl-CoA carboxylase PASTICCINO3 and the 3-hydroxy acyl-CoA dehydratase PASTICCINO2 revealed that VLCFAs are important for cell proliferation and tissue patterning. Here, we show that the immunophilin PASTICCINO1 (PAS1) is also required for VLCFA synthesis. Impairment of PAS1 function results in reduction of VLCFA levels that particularly affects the composition of sphingolipids, known to be important for cell polarity in animals. Moreover, PAS1 associates with several enzymes of the VLCFA elongase complex in the endoplasmic reticulum. The pas1 mutants are deficient in lateral root formation and are characterized by an abnormal patterning of the embryo apex, which leads to defective cotyledon organogenesis. Our data indicate that in both tissues, defective organogenesis is associated with the mistargeting of the auxin efflux carrier PIN FORMED1 in specific cells, resulting in local alteration of polar auxin distribution. Furthermore, we show that exogenous VLCFAs rescue lateral root organogenesis and polar auxin distribution, indicating their direct involvement in these processes. Based on these data, we propose that PAS1 acts as a molecular scaffold for the fatty acid elongase complex in the endoplasmic reticulum and that the resulting VLCFAs are required for polar auxin transport and tissue patterning during plant development.
Figures
References
-
- Aida M., Vernoux T., Furutani M., Traas J., Tasaka M. (2002). Roles of PIN-FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo. Development 129: 3965–3974 - PubMed
-
- Baud S., Guyon V., Kronenberger J., Wuillème S., Miquel M., Caboche M., Lepiniec L., Rochat C. (2003). Multifonctional acetyl-CoA carboxylase 1 is essential for very long chain fatty acids elongation and embryo development in Arabidopsis. Plant J. 33: 75–86 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
