

Comparative Analyses of Transcriptional Profiles in Mouse Organs Using a Pneumonic Plague Model after Infection with Wild-Type Yersinia pestis CO92 and Its Braun Lipoprotein Mutant
- PMID: 20145715
- PMCID: PMC2817383
- DOI: 10.1155/2009/914762
Comparative Analyses of Transcriptional Profiles in Mouse Organs Using a Pneumonic Plague Model after Infection with Wild-Type Yersinia pestis CO92 and Its Braun Lipoprotein Mutant
Abstract
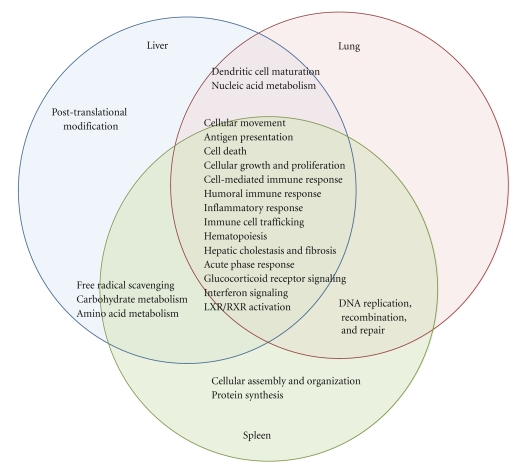
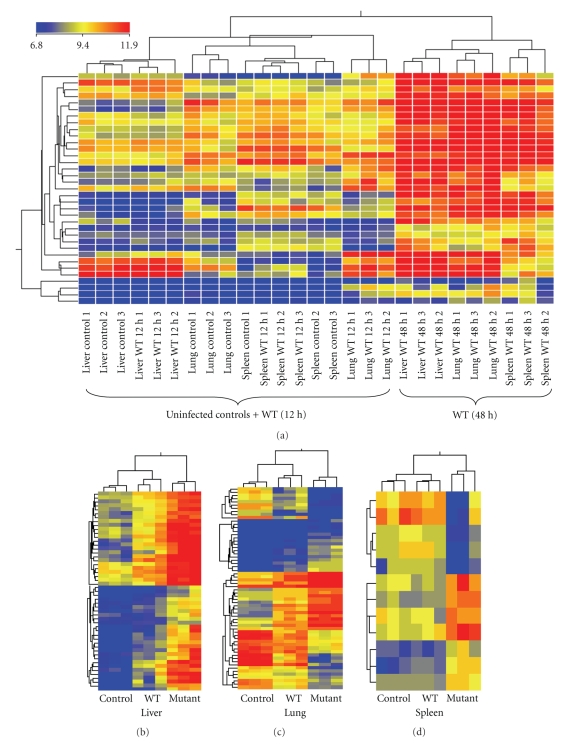
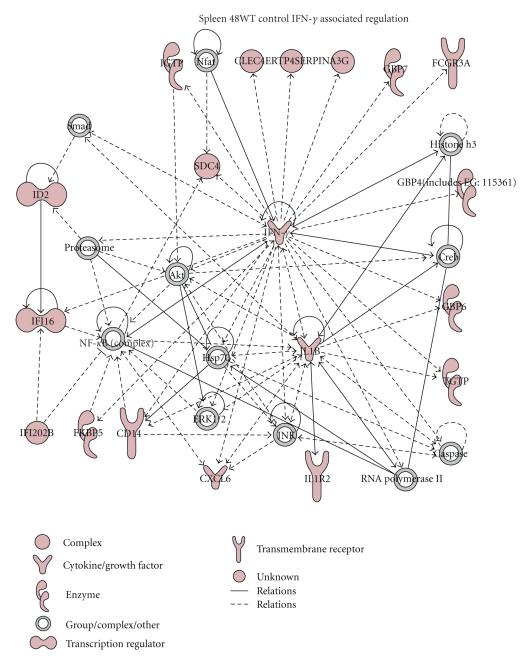
We employed Murine GeneChips to delineate the global transcriptional profiles of the livers, lungs, and spleens in a mouse pneumonic plague infection model with wild-type (WT) Y. pestis CO92 and its Braun lipoprotein (Deltalpp) mutant with reduced virulence. These organs showed differential transcriptional responses to infection with WT Y. pestis, but the overall host functional processes affected were similar across all three tissues. Gene expression alterations were found in inflammation, cytokine signaling, and apoptotic cell death-associated genes. Comparison of WT and Deltalpp mutant-infected mice indicated significant overlap in lipopolysaccharide- (LPS-) associated gene expression, but the absence of Lpp perturbed host cell signaling at critical regulatory junctions resulting in altered immune response and possibly host cell apoptosis. We generated a putative signaling pathway including major inflammatory components that could account for the synergistic action of LPS and Lpp and provided the mechanistic basis of attenuation caused by deletion of the lpp gene from Y. pestis in a mouse model of pneumonic plague.
Figures
Similar articles
-
Further characterization of a highly attenuated Yersinia pestis CO92 mutant deleted for the genes encoding Braun lipoprotein and plasminogen activator protease in murine alveolar and primary human macrophages.Microb Pathog. 2015 Mar;80:27-38. doi: 10.1016/j.micpath.2015.02.005. Epub 2015 Feb 16. Microb Pathog. 2015. PMID: 25697665 Free PMC article.
-
Deletion of Braun lipoprotein gene (lpp) and curing of plasmid pPCP1 dramatically alter the virulence of Yersinia pestis CO92 in a mouse model of pneumonic plague.Microbiology (Reading). 2009 Oct;155(Pt 10):3247-3259. doi: 10.1099/mic.0.029124-0. Epub 2009 Jul 9. Microbiology (Reading). 2009. PMID: 19589835 Free PMC article.
-
Braun lipoprotein (Lpp) contributes to virulence of yersiniae: potential role of Lpp in inducing bubonic and pneumonic plague.Infect Immun. 2008 Apr;76(4):1390-409. doi: 10.1128/IAI.01529-07. Epub 2008 Jan 28. Infect Immun. 2008. PMID: 18227160 Free PMC article.
-
Yersinia pestis and pneumonic plague: Insight into how a lethal pathogen interfaces with innate immune populations in the lung to cause severe disease.Cell Immunol. 2024 Sep-Oct;403-404:104856. doi: 10.1016/j.cellimm.2024.104856. Epub 2024 Jul 10. Cell Immunol. 2024. PMID: 39002222 Review.
-
Pneumonic Plague: The Darker Side of Yersinia pestis.Trends Microbiol. 2016 Mar;24(3):190-197. doi: 10.1016/j.tim.2015.11.008. Epub 2015 Dec 14. Trends Microbiol. 2016. PMID: 26698952 Review.
Cited by
-
Identification of modulators of the nuclear receptor peroxisome proliferator-activated receptor α (PPARα) in a mouse liver gene expression compendium.PLoS One. 2015 Feb 17;10(2):e0112655. doi: 10.1371/journal.pone.0112655. eCollection 2015. PLoS One. 2015. PMID: 25689681 Free PMC article.
-
Further characterization of a highly attenuated Yersinia pestis CO92 mutant deleted for the genes encoding Braun lipoprotein and plasminogen activator protease in murine alveolar and primary human macrophages.Microb Pathog. 2015 Mar;80:27-38. doi: 10.1016/j.micpath.2015.02.005. Epub 2015 Feb 16. Microb Pathog. 2015. PMID: 25697665 Free PMC article.
-
Temporal Progression of Pneumonic Plague in Blood of Nonhuman Primate: A Transcriptomic Analysis.PLoS One. 2016 Mar 22;11(3):e0151788. doi: 10.1371/journal.pone.0151788. eCollection 2016. PLoS One. 2016. PMID: 27003632 Free PMC article.
-
Omics strategies for revealing Yersinia pestis virulence.Front Cell Infect Microbiol. 2012 Dec 13;2:157. doi: 10.3389/fcimb.2012.00157. eCollection 2012. Front Cell Infect Microbiol. 2012. PMID: 23248778 Free PMC article. Review.
-
Comparative Global Gene Expression Profiles of Wild-Type Yersinia pestis CO92 and Its Braun Lipoprotein Mutant at Flea and Human Body Temperatures.Comp Funct Genomics. 2010;2010:342168. doi: 10.1155/2010/342168. Epub 2010 May 19. Comp Funct Genomics. 2010. PMID: 20508723 Free PMC article.
References
-
- Inglesby TV, Dennis DT, Henderson DA, et al. Plague as a biological weapon: medical and public health management. Journal of the American Medical Association. 2000;283(17):2281–2290. - PubMed
-
- Adkins I, Köberle M, Gröbner S, Bohn E, Autenrieth IB, Borgmann S. Yersinia outer proteins E, H, P, and T differentially target the cytoskeleton and inhibit phagocytic capacity of dendritic cells. International Journal of Medical Microbiology. 2007;297(4):235–244. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases