

P2X7, NMDA and BDNF receptors converge on GSK3 phosphorylation and cooperate to promote survival in cerebellar granule neurons
- PMID: 20146080
- PMCID: PMC2858808
- DOI: 10.1007/s00018-010-0278-x
P2X7, NMDA and BDNF receptors converge on GSK3 phosphorylation and cooperate to promote survival in cerebellar granule neurons
Abstract
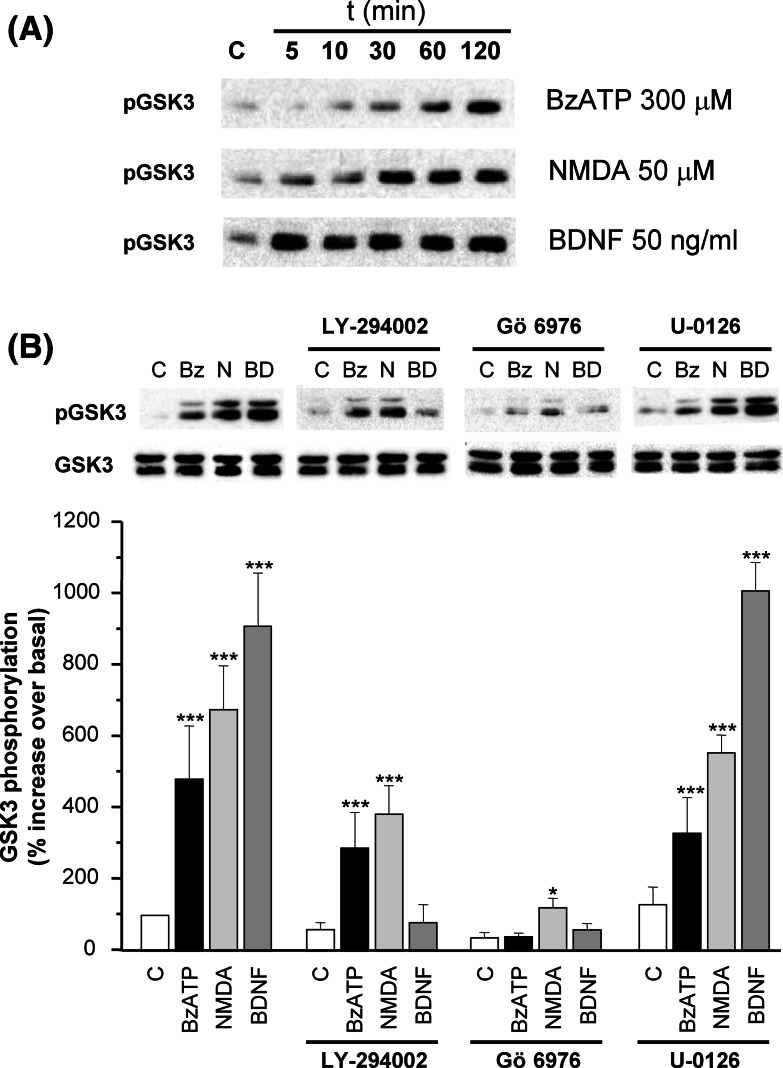
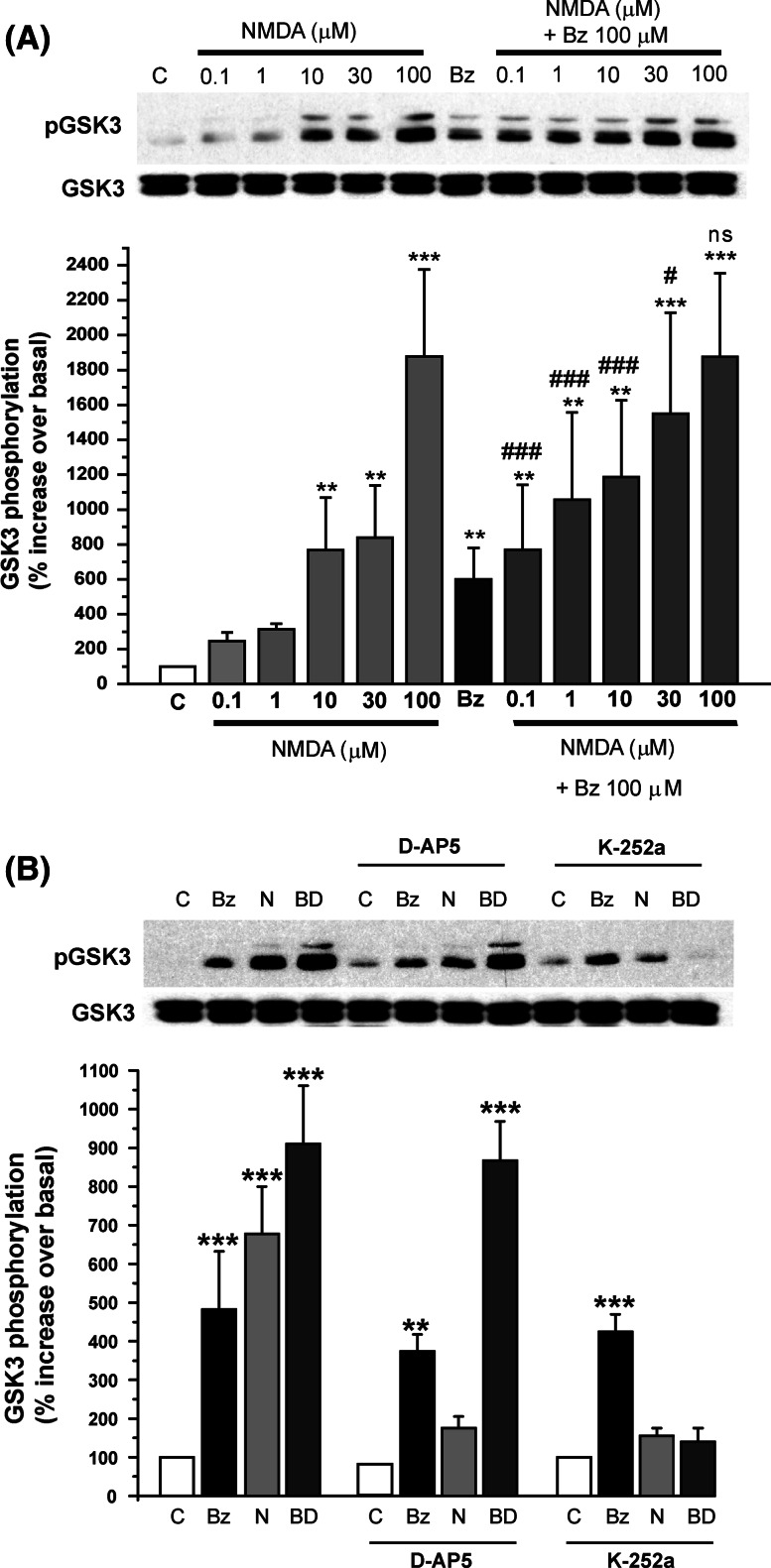
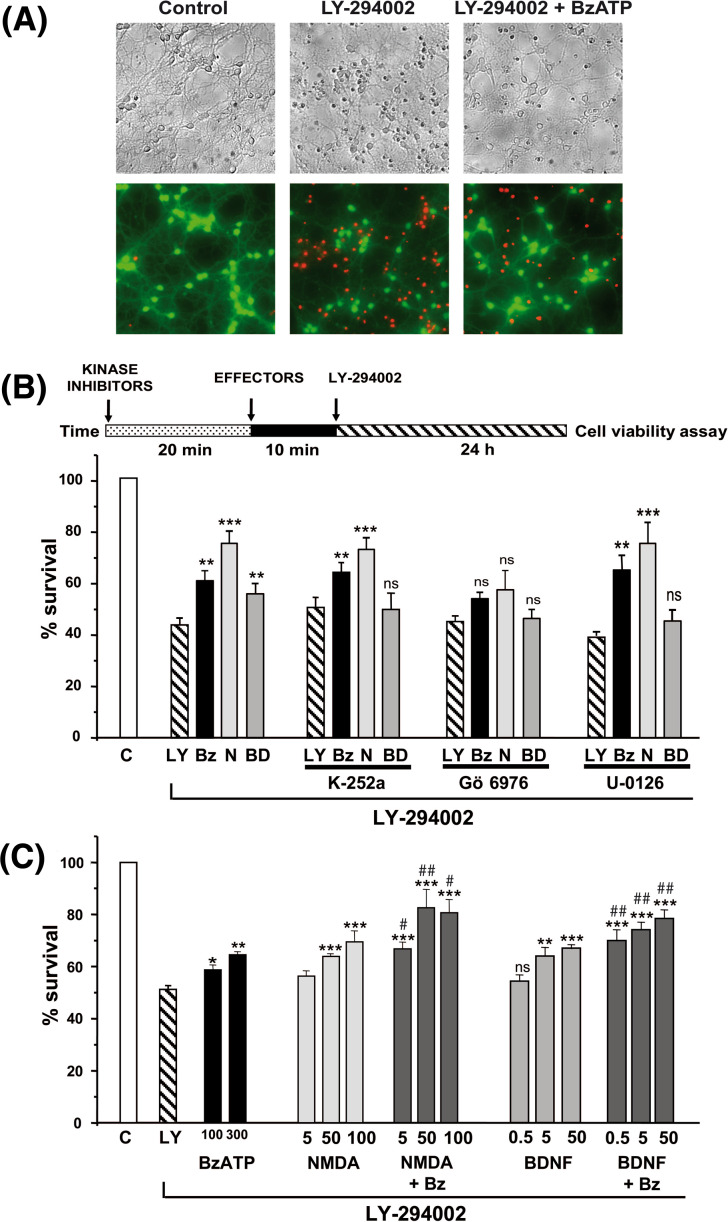
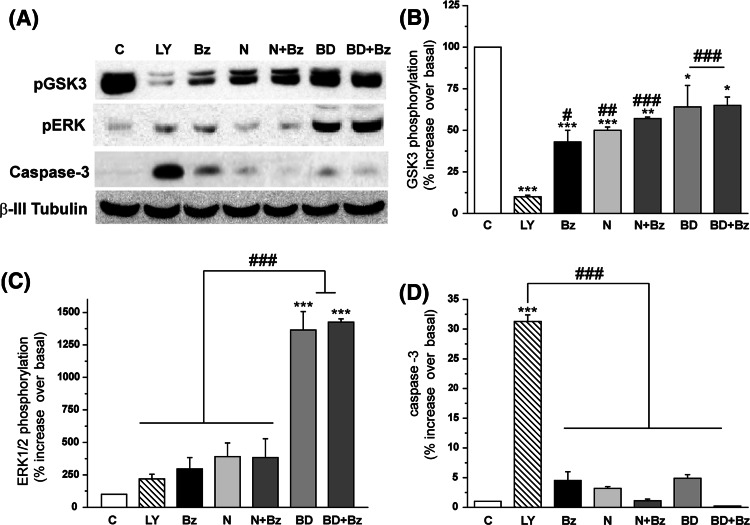
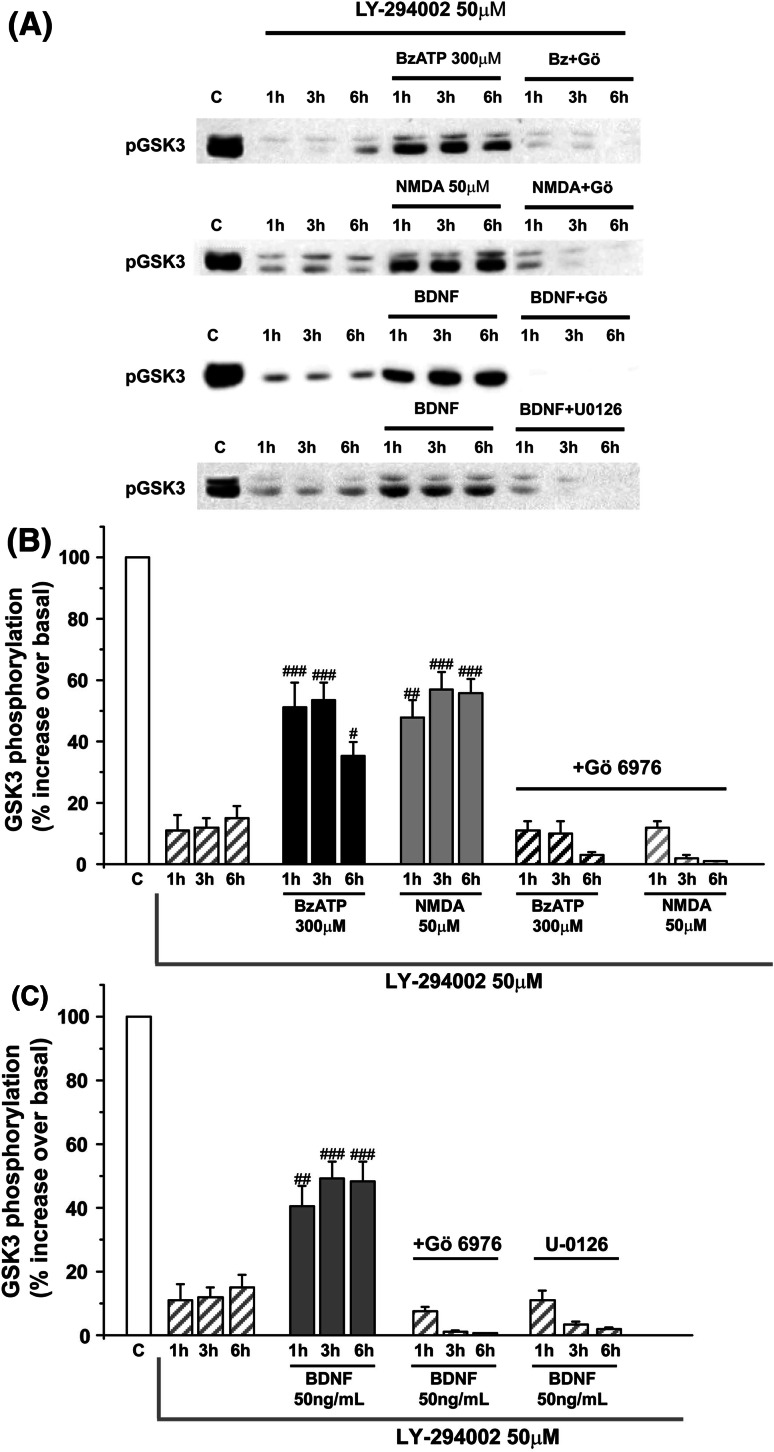
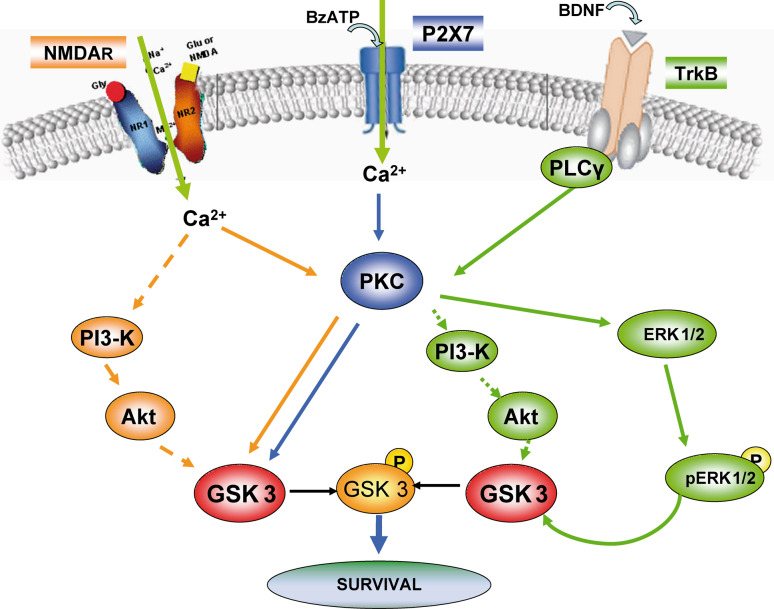
Glycogen synthase kinase-3 (GSK3) is a key player in the regulation of neuronal survival. Herein, we report evidence of an interaction between P2X7 receptors with NMDA and BDNF receptors at the level of GSK3 signalling and neuroprotection. The activation of these receptors in granule neurons led to a sustained pattern of GSK3 phosphorylation that was mainly PKC-dependent. BDNF was the most potent at inducing GSK3 phosphorylation, which was also dependent on PI3K. The P2X7 agonist, BzATP, exhibited additive effects with both NMDA and BDNF to rescue granule neurons from cell death induced by PI3K inhibition. This survival effect was mediated by the PKC-dependent GSK3 pathway. In addition, ERK1/2 proteins were also involved in BDNF protective effect. These results show the function of ATP in amplifying neuroprotective actions of glutamate and neurotrophins, and support the role of GSK3 as an important convergence point for these survival promoting factors in granule neurons.
Figures
Similar articles
-
P2X7 nucleotide receptor is coupled to GSK-3 inhibition and neuroprotection in cerebellar granule neurons.Neurotox Res. 2009 Apr;15(3):193-204. doi: 10.1007/s12640-009-9020-6. Epub 2009 Feb 24. Neurotox Res. 2009. PMID: 19384592
-
ERK1/2 activation is involved in the neuroprotective action of P2Y13 and P2X7 receptors against glutamate excitotoxicity in cerebellar granule neurons.Neuropharmacology. 2011 Dec;61(8):1210-21. doi: 10.1016/j.neuropharm.2011.07.010. Epub 2011 Jul 28. Neuropharmacology. 2011. PMID: 21798274
-
Co-activation of the phosphatidylinositol-3-kinase/Akt signaling pathway by N-methyl-D-aspartate and TrkB receptors in cerebellar granule cell neurons.Amino Acids. 2002;23(1-3):11-7. doi: 10.1007/s00726-001-0103-9. Amino Acids. 2002. PMID: 12373512
-
N-methyl-D-aspartate and TrkB receptor activation in cerebellar granule cells: an in vitro model of preconditioning to stimulate intrinsic survival pathways in neurons.Ann N Y Acad Sci. 2003 May;993:134-45; discussion 159-60. doi: 10.1111/j.1749-6632.2003.tb07522.x. Ann N Y Acad Sci. 2003. PMID: 12853306 Free PMC article. Review.
-
P2 receptor interaction and signalling cascades in neuroprotection.Brain Res Bull. 2019 Sep;151:74-83. doi: 10.1016/j.brainresbull.2018.12.012. Epub 2018 Dec 26. Brain Res Bull. 2019. PMID: 30593879 Review.
Cited by
-
Brain-derived neurotrophic factor in Alzheimer's disease and its pharmaceutical potential.Transl Neurodegener. 2022 Jan 28;11(1):4. doi: 10.1186/s40035-022-00279-0. Transl Neurodegener. 2022. PMID: 35090576 Free PMC article. Review.
-
Role of P2X7 and P2Y2 receptors on α-secretase-dependent APP processing: Control of amyloid plaques formation "in vivo" by P2X7 receptor.Comput Struct Biotechnol J. 2015 Mar 11;13:176-81. doi: 10.1016/j.csbj.2015.02.005. eCollection 2015. Comput Struct Biotechnol J. 2015. PMID: 25848496 Free PMC article. Review.
-
Effects of Treadmill Exercise on the Recovery of Dopaminergic Neuron Loss and Muscle Atrophy in the 6-OHDA Lesioned Parkinson's Disease Rat Model.Korean J Physiol Pharmacol. 2012 Oct;16(5):305-12. doi: 10.4196/kjpp.2012.16.5.305. Epub 2012 Oct 18. Korean J Physiol Pharmacol. 2012. PMID: 23129977 Free PMC article.
-
P2X7 nucleotide receptor signaling potentiates the Wnt/β-catenin pathway in cells of the osteoblast lineage.Purinergic Signal. 2016 Sep;12(3):509-20. doi: 10.1007/s11302-016-9517-4. Epub 2016 May 21. Purinergic Signal. 2016. PMID: 27206526 Free PMC article.
-
Alternative Pharmacological Strategies for the Treatment of Alzheimer's Disease: Focus on Neuromodulator Function.Biomedicines. 2022 Nov 28;10(12):3064. doi: 10.3390/biomedicines10123064. Biomedicines. 2022. PMID: 36551821 Free PMC article. Review.
References
-
- North RA. Molecular physiology of P2X receptors. Physiol Rev. 2002;82:1013–1067. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
