

Parametric manipulation of the conflict signal and control-state adaptation
- PMID: 20146615
- PMCID: PMC3056266
- DOI: 10.1162/jocn.2010.21458
Parametric manipulation of the conflict signal and control-state adaptation
Abstract
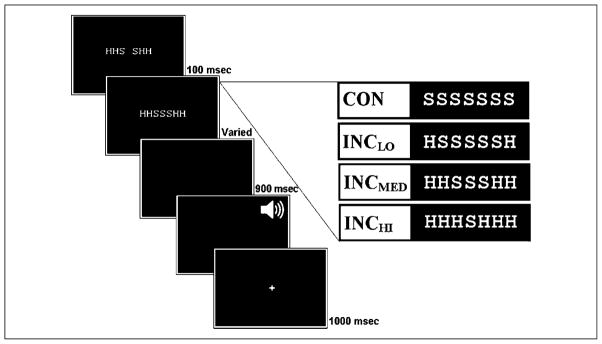
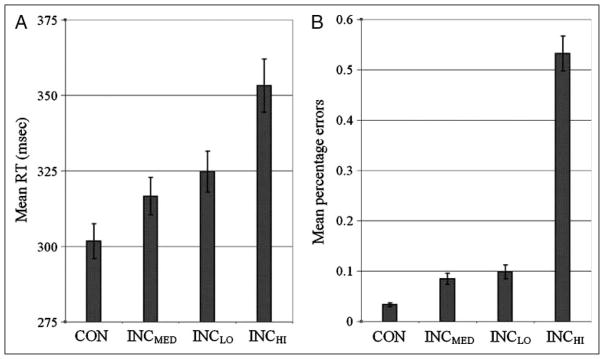
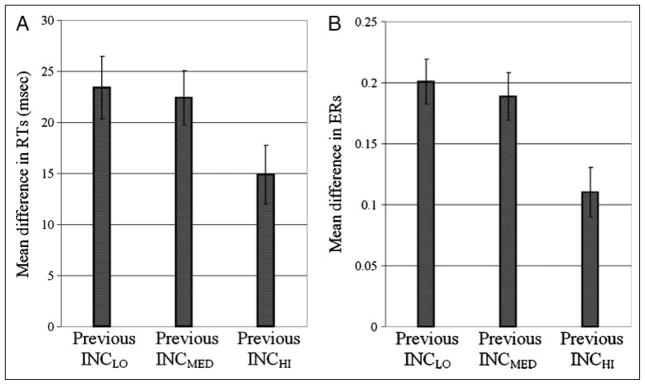
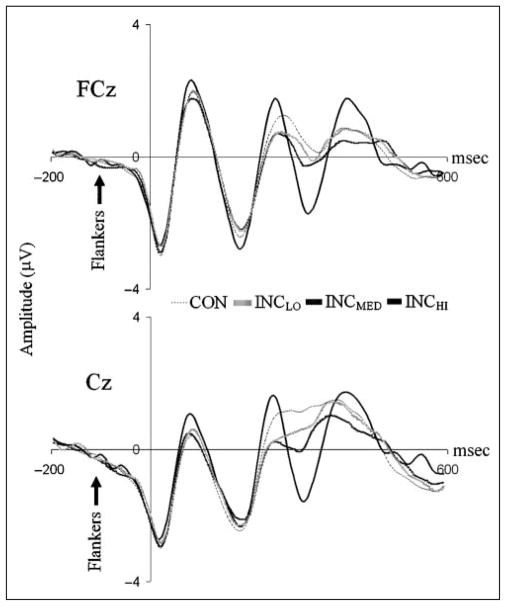
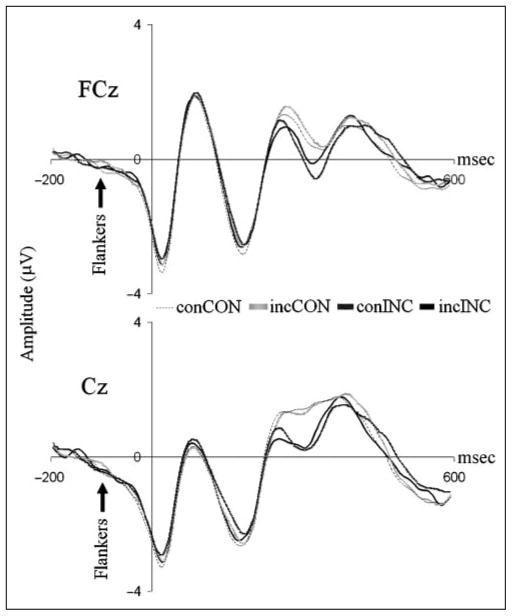
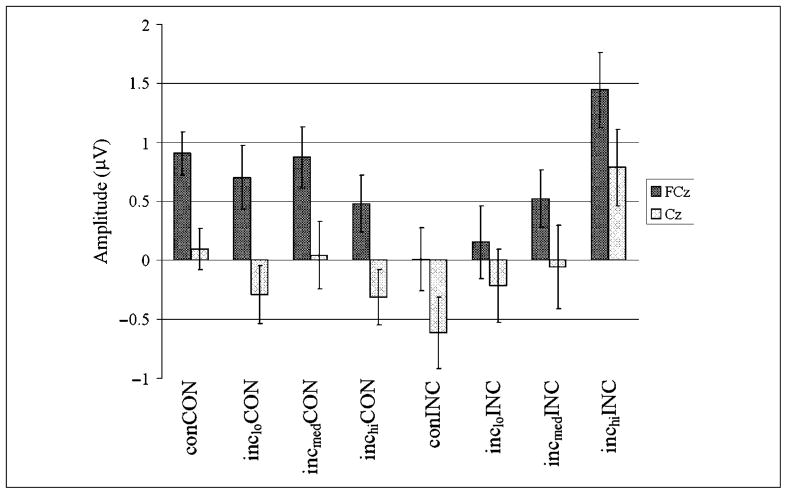
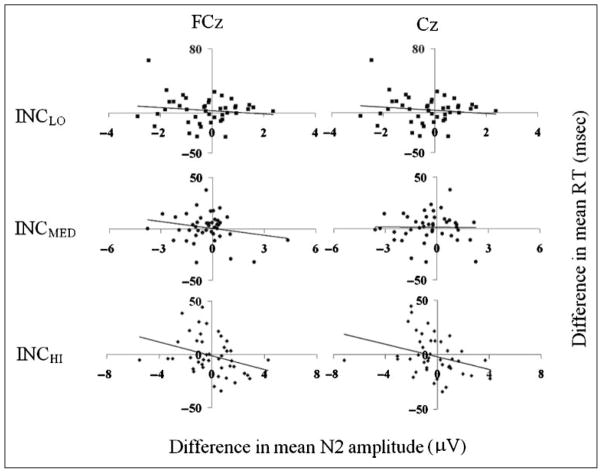
Mechanisms by which the brain monitors and modulates performance are an important focus of recent research. The conflict-monitoring hypothesis posits that the ACC detects conflict between competing response pathways which, in turn, signals for enhanced control. The N2, an ERP component that has been localized to ACC, has been observed after high conflict stimuli. As a candidate index of the conflict signal, the N2 would be expected to be sensitive to the degree of response conflict present, a factor that depends on both the features of external stimuli and the internal control state. In the present study, we sought to explore the relationship between N2 amplitude and these variables through use of a modified Eriksen flankers task in which target-distracter compatibility was parametrically varied. We hypothesized that greater target-distracter incompatibility would result in higher levels of response conflict, as indexed by both behavior and the N2 component. Consistent with this prediction, there were parametric degradations in behavioral performance and increases in N2 amplitudes with increasing incompatibility. Further, increasingly incompatible stimuli led to the predicted parametric increases in control on subsequent incompatible trials as evidenced by enhanced performance and reduced N2 amplitudes. These findings suggest that the N2 component and associated behavioral performance are finely sensitive to the degree of response conflict present and to the control adjustments that result from modulations in conflict.
Figures
Similar articles
-
Performance monitoring following conflict: internal adjustments in cognitive control?Neuropsychologia. 2012 Feb;50(3):426-33. doi: 10.1016/j.neuropsychologia.2011.12.021. Epub 2012 Jan 3. Neuropsychologia. 2012. PMID: 22234168
-
Conflict adaptation and sequential trial effects: support for the conflict monitoring theory.Neuropsychologia. 2011 Jun;49(7):1953-61. doi: 10.1016/j.neuropsychologia.2011.03.023. Epub 2011 Mar 22. Neuropsychologia. 2011. PMID: 21435347
-
Cognitive conflict adaptation in generalized anxiety disorder.Biol Psychol. 2013 Oct;94(2):408-18. doi: 10.1016/j.biopsycho.2013.08.006. Epub 2013 Sep 1. Biol Psychol. 2013. PMID: 24005064
-
Making sense of all the conflict: a theoretical review and critique of conflict-related ERPs.Int J Psychophysiol. 2014 Sep;93(3):283-97. doi: 10.1016/j.ijpsycho.2014.06.007. Epub 2014 Jun 17. Int J Psychophysiol. 2014. PMID: 24950132 Review.
-
The anterior cingulate as a conflict monitor: fMRI and ERP studies.Physiol Behav. 2002 Dec;77(4-5):477-82. doi: 10.1016/s0031-9384(02)00930-7. Physiol Behav. 2002. PMID: 12526986 Review.
Cited by
-
Cognitive control adjustments are dependent on the level of conflict.Sci Rep. 2024 Feb 7;14(1):3116. doi: 10.1038/s41598-024-53090-4. Sci Rep. 2024. PMID: 38326342 Free PMC article.
-
Exploring behavioral adjustments of proportion congruency manipulations in an Eriksen flanker task with visual and auditory distractor modalities.Mem Cognit. 2024 Jan;52(1):91-114. doi: 10.3758/s13421-023-01447-x. Epub 2023 Aug 7. Mem Cognit. 2024. PMID: 37548866 Free PMC article.
-
The influence of approach-avoidance motivational orientation on conflict adaptation.Cogn Affect Behav Neurosci. 2014 Jun;14(2):548-60. doi: 10.3758/s13415-014-0295-6. Cogn Affect Behav Neurosci. 2014. PMID: 24841080
-
Evidence for a role for the dorsal anterior cingulate cortex in disengaging from an incorrect action.PLoS One. 2014 Jun 26;9(6):e101126. doi: 10.1371/journal.pone.0101126. eCollection 2014. PLoS One. 2014. PMID: 24968256 Free PMC article.
-
Event-related potentials reflect impaired temporal interval learning following haloperidol administration.Psychopharmacology (Berl). 2017 Sep;234(17):2545-2562. doi: 10.1007/s00213-017-4645-2. Epub 2017 Jun 10. Psychopharmacology (Berl). 2017. PMID: 28601965 Clinical Trial.
References
-
- Allport DA, Styles E, Hsieh S. Switching intentional set: Exploring the dynamic control of tasks. In: Umilta C, Moscovitch M, editors. Attention and Performance XV: Conscious and nonconscious information processing. Cambridge, MA: MIT Press; 1994. pp. 421–452.
-
- Badre D, Wagner AD. Selection, integration, and conflict monitoring; assessing the nature and generality of prefrontal cognitive control mechanisms. Neuron. 2004;41:473–487. - PubMed
-
- Beste C, Saft C, Andrich J, Gold R, Falkenstein M. Stimulus-response compatibility in Huntington’s disease: A cognitive-neurophysiological analysis. Journal of Neurophysiology. 2008;99:1213–1223. - PubMed
-
- Botvinick MM. Conflict monitoring and decision making: Reconciling two perspectives on anterior cingulate function. Cognitive, Affective & Behavioral Neuroscience. 2007;7:356–366. - PubMed
-
- Botvinick M, Braver TS, Barch DM, Carter CS, Cohen JD. Conflict monitoring and cognitive control. Psychological Review. 2001;108:624–652. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials