

Review
doi: 10.1242/dev.038695.
Making a difference together: reciprocal interactions in C. elegans and zebrafish asymmetric neural development
Affiliations
- PMID: 20147373
- PMCID: PMC2827681
- DOI: 10.1242/dev.038695
Item in Clipboard
Review
Making a difference together: reciprocal interactions in C. elegans and zebrafish asymmetric neural development
Development.
2010 Mar.
Abstract
Brain asymmetries are thought to increase neural processing capacity and to prevent interhemispheric conflict. In order to develop asymmetrically, neurons must be specified along the left-right axis, assigned left-side versus right-side identities and differentiate appropriately. In C. elegans and zebrafish, the cellular and molecular mechanisms that lead to neural asymmetries have recently come to light. Here, we consider recent insights into the mechanisms involved in asymmetrical neural development in these two species. Although the molecular details are divergent, both organisms use iterative cell-cell communication to establish left-right neuronal identity.
Figures
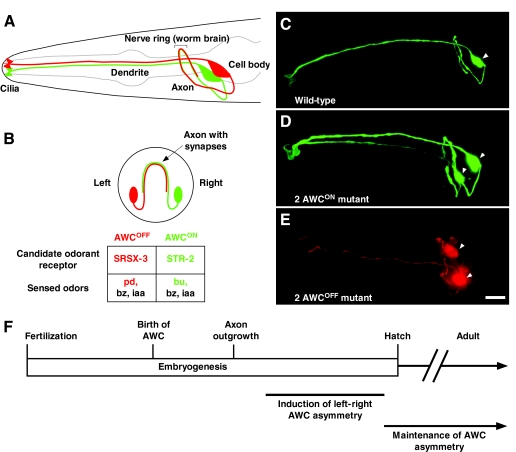
Left-right AWC neuronal asymmetry in C. elegans. (A,B) Amphid wing `C' (AWC) cell anatomy. Lateral (A) and transverse (B) views of a wild-type C. elegans head showing two bilateral AWC neurons. One AWC cell that expresses a GFP-tagged transgene of the candidate odorant receptor gene str-2 (str-2::GFP) (AWCON; right) is shown in green; the other AWC, with no str-2::GFP expression (AWCOFF; left), is shown in red. Within a population of worms, the left-right AWC asymmetry is stochastic, meaning that 50% of the animals display str-2 expression on the right, whereas the other 50% display str-2 expression on the left. The AWCON cell expresses the STR-2 candidate odorant receptor and senses butanone (bu), benzaldehyde (bz) and isoamyl alcohol (iaa). The AWCOFF cell expresses the SRSX-3 candidate odorant receptor and senses pentanedione (pd), bz and iaa (Bargmann et al., 1993; Wes and Bargmann, 2001). (C-E) Projections of micrograph stacks of wild-type and mutant animals expressing str-2::GFP (pseudo-colored green for GFP, red for DsRed). (C) Wild-type animals express str-2::GFP in one AWC neuron. (D) 2 AWCON animals express str-2::GFP in both AWC neurons. (E) Bilateral expression of the AWC marker transgene odr-1::DsRed in a 2 AWCOFF mutant showing the presence of both AWCs without str-2::GFP expression. (F) Time-line of developmental events in left-right AWC asymmetry. The AWC neurons are born at 300 minutes and their axons extend at 450 minutes after fertilization. AWC asymmetry is established during late embryogenesis and is maintained throughout the life of the animal. Arrowheads indicate the cell body of AWC neurons. Ventral is at the bottom in A-E, and anterior is to the left in A,C-E. Scale bar: 10 μm.
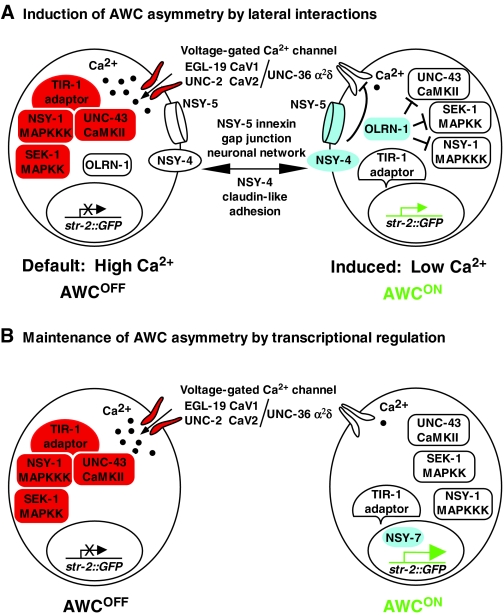
Left-right AWC asymmetry is controlled by calcium. (A) Induction of left-right AWC asymmetry by lateral interactions. The AWCOFF default state is specified by calcium-MAPK signaling. Before cell-cell communication is initiated, AWC cells undergo a calcium influx through voltage-gated calcium channels. Calcium signaling activates the CaMKII-MAPK pathway, mediated by the TIR-1 adaptor protein, resulting in the inactivity of the str-2 promoter and in the AWCOFF state in both AWC cells. After axonal outgrowth, the NSY-5 innexin gap junction neuronal network and the NSY-4 claudin-like protein activate the Raw repeat protein OLRN-1 to antagonize the calcium-mediated signaling pathway in the AWC that will induce str-2 promoter activity and become AWCON. NSY-4 and NSY-5 are more active in the future AWCON cell than the future AWCOFF cell (see Fig. 3 and text for details). (B) Maintenance of left-right AWC asymmetry by transcriptional regulation. The NSY-7 transcription factor maintains str-2 promoter activity in the induced AWCON cell and represses AWCOFF genes, such as srsx-3 (not shown).
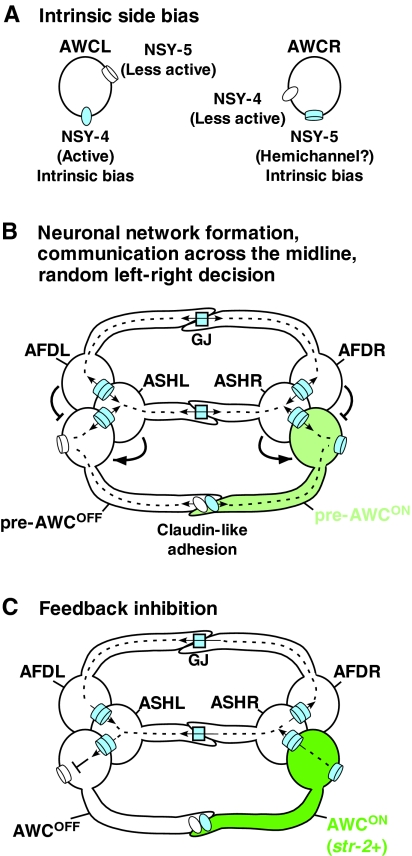
Model of a neuronal signaling network for AWC asymmetry. (A) Intrinsic side bias of NSY-4 claudin-like protein and NSY-5 innexin gap juction protein in AWC neurons. NSY-4 activity has a bias toward the left AWC cell (AWCL), whereas NSY-5 activity, perhaps present as hemichannels, has a bias toward the right AWC cell (AWCR). (B) Random left-right AWC asymmetry decisions via neuronal network formation and communication across the midline. Gap junctions (GJ) and claudin-like adhesions connect bilateral AWC neurons and their neighboring neurons, including AFD and ASH neurons, in a network. Gap junctions mediate direct contact between the cell bodies of the respective neurons on each side, whereas axonal contacts through gap junctions and claudin-like adhesions mediate signaling across the midline between left and right neurons. AFD cells have a negative influence on the ipsilateral cell becoming AWCON, and ASH cells have a positive influence on the ipsilateral cell becoming AWCON. Both NSY-4 and NSY-5 are required to induce AWCON and the relative strengths of NSY-4 and NSY-5 activity between the two AWC cells specify random left-right asymmetry. The pre-AWCON cell indicates the AWC cell that turns on str-2 expression. (C) Feedback inhibition of NSY-4 and NSY-5 across the midline from the AWCON cell to the contralateral AWCOFF cell coordinates and strengthens str-2 gene expression in only one AWC cell.
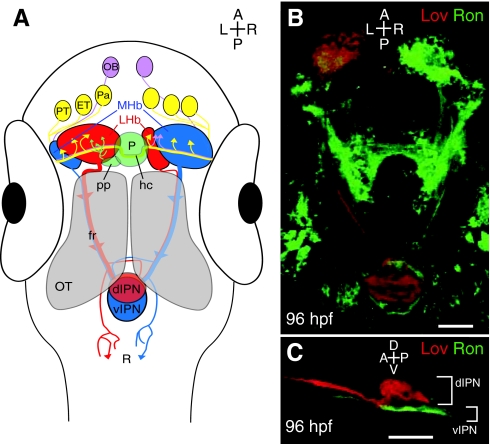
The asymmetric zebrafish habenular nuclei are central to the dorsal diencephalic conduction pathway. (A) The dorsal diencephalic conduction system. The ratio of lateral (red) to medial (blue) subnuclei is greater in the left habenula, but both sides receive input from both ipsilateral and contralateral neurons of the pallium, the eminentia thalami and the posterior tuberculum. The parapineal only sends projections to the left habenula, whereas the right habenula receives additional afferents from both olfactory bulbs. Habenular efferents course ventrally to the optic tecta in converging bundles known as the fasciculus retroflexi. Axons originating in the medial subnuclei innervate the ventral region of the interpeduncular nucleus, whereas those of lateral origin can target the dorsal interpeduncular nucleus. Some habenular axons also contact the raphe, just posterior to the interpeduncular nucleus. (B) Dorsal perspective confocal z-projection of Leftover (Lov)-positive (Lov+; red) and Right on (Ron)-positive (Ron+; green) neurons coursing from the habenular nuclei to the interpeduncular nucleus via the fasciculus retroflexi. (C) Lateral view of Lov+ and Ron+ axons, segregated into the dorsal and ventral domains of the interpeduncular nucleus. dIPN, dorsal interpeduncular nucleus; ET, eminentia thalami; fr, fasciculus retroflexus; hc, habenular commissure; LHb, lateral habenular subnuclei; MHb, medial habenular subnuclei; OB, olfactory bulb; OT, optic tectum; P, pineal organ; Pa, pallium; pp, parapineal organ; PT, posterior tuberculum; R, raphe nucleus; vIPN, ventral interpeduncular nucleus. Scale bars: 50 μm.
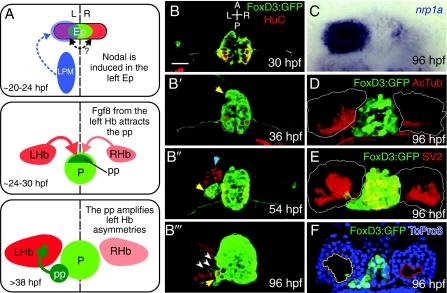
Asymmetric development of the zebrafish epithalamus. The zebrafish epithalamus begins as a symmetrical cell population and develops a suite of asymmetries. (A) Model of asymmetric epithalamic development. First, around 20 hours post-fertilization (hpf), Nodal expression (blue) in the left lateral plate mesoderm (LPM) activates Nodal expression in the left epithalamus (Ep). There might be communication across the midline in the epithalamus (double-headed arrow; see text for discussion). At around 24 hpf, a slight left-sided bias of Fgf8 from the habenulae (Hb) attracts the parapineal (pp). By 38 hpf, the parapineal induces the elaboration of asymmetries only in the left habenula. (B-B″′) Timecourse of epithalamic development. Tg(foxd3:GFP) (green) marks the pineal and parapineal, and mature neurons are marked with antibodies against HuC (red). (B) At 30 hpf, the pineal is a symmetrical group of neurons centered on the midline. (B′) By 36 hpf, the parapineal (yellow arrow) begins to migrate away from the left anterior border of the pineal. (B″) At 54 hpf, the parapineal (yellow arrow) is a distinct accessory organ, and more neurons in the left habenula (blue arrow) have differentiated (i.e. are HuC-positive) than in the right. (B″′) The 96 hpf epithalamus is robustly lateralized, with parapineal axons (white arrows) contacting only the left habenula, which is made up of many more HuC-positive neurons than the right. (C-F) Asymmetrical features of the habenular nuclei at 96 hpf. (C) The transcript of the gene encoding the semaphorin co-receptor neuropilin1a (nrp1a) (blue) is expressed predominantly on the left side in the lateral habenular subnucleus. (D) Antibodies against acetylated tubulin (red) reveal all neuronal processes and show a greater density of neuropil in the left habenula (habenular borders outlined in white). (E) Antibodies against the presynaptic component SV2 (red) demonstrate a greater density of synaptic contacts in the left habenula. (F) An optical slice through the left and right habenulae at the level of the parapineal stained with the nuclear marker To-Pro3 (blue) demonstrates that the soma-free core of the left habenula (yellow outline) is larger than that in the right (red outline) and is invaded by parapineal axons. Scale bar: 50 μm.
Similar articles
-
Gap junctions provide new links in left-right patterning.Cell. 2007 May 18;129(4):645-7. doi: 10.1016/j.cell.2007.05.005. Cell. 2007. PMID: 17512395 Review.
-
Left-right asymmetry: making the right decision early.Curr Biol. 2006 Dec 19;16(24):R1039-42. doi: 10.1016/j.cub.2006.11.009. Curr Biol. 2006. PMID: 17174913
-
Stochastic left-right neuronal asymmetry in Caenorhabditis elegans.Philos Trans R Soc Lond B Biol Sci. 2016 Dec 19;371(1710):20150407. doi: 10.1098/rstb.2015.0407. Philos Trans R Soc Lond B Biol Sci. 2016. PMID: 27821536 Free PMC article. Review.
-
Asymmetric neural development in the Caenorhabditis elegans olfactory system.Genesis. 2014 Jun;52(6):544-54. doi: 10.1002/dvg.22744. Epub 2014 Feb 7. Genesis. 2014. PMID: 24478264 Free PMC article. Review.
-
An innexin-dependent cell network establishes left-right neuronal asymmetry in C. elegans.Cell. 2007 May 18;129(4):787-99. doi: 10.1016/j.cell.2007.02.052. Cell. 2007. PMID: 17512411
Cited by
-
Intercellular calcium signaling in a gap junction-coupled cell network establishes asymmetric neuronal fates in C. elegans.Development. 2012 Nov;139(22):4191-201. doi: 10.1242/dev.083428. Development. 2012. PMID: 23093425 Free PMC article.
-
FGF signaling is required for brain left-right asymmetry and brain midline formation.Dev Biol. 2014 Feb 1;386(1):123-34. doi: 10.1016/j.ydbio.2013.11.020. Epub 2013 Dec 12. Dev Biol. 2014. PMID: 24333178 Free PMC article.
-
Microtubule-based localization of a synaptic calcium-signaling complex is required for left-right neuronal asymmetry in C. elegans.Development. 2011 Aug;138(16):3509-18. doi: 10.1242/dev.069740. Epub 2011 Jul 19. Development. 2011. PMID: 21771813 Free PMC article.
-
Embryonic Ethanol Exposure Affects the Early Development, Migration, and Location of Hypocretin/Orexin Neurons in Zebrafish.Alcohol Clin Exp Res. 2019 Aug;43(8):1702-1713. doi: 10.1111/acer.14126. Epub 2019 Jul 6. Alcohol Clin Exp Res. 2019. PMID: 31206717 Free PMC article.
-
Fgf signaling governs cell fate in the zebrafish pineal complex.Development. 2013 Jan 15;140(2):323-32. doi: 10.1242/dev.083709. Development. 2013. PMID: 23250206 Free PMC article.
References
-
- Ahmad N., Long S., Rebagliati M. (2004). A southpaw joins the roster: the role of the zebrafish nodal-related gene southpaw in cardiac LR asymmetry. Trends Cardiovasc. Med. 14, 43-49 - PubMed
-
- Aizawa H., Goto M., Sato T., Okamoto H. (2007). Temporally regulated asymmetric neurogenesis causes left-right difference in the zebrafish habenular structures. Dev. Cell 12, 87-98 - PubMed
-
- Amunts K., Schlaug G., Schleicher A., Steinmetz H., Dabringhaus A., Roland P. E., Zilles K. (1996). Asymmetry in the human motor cortex and handedness. Neuroimage 4, 216-222 - PubMed
-
- Asai T., Sugimori E., Tanno Y. (2009). Schizotypal personality traits and atypical lateralization in motor and language functions. Brain Cogn. 71, 26-37 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
