

Stimulation of HIV-1 replication in immature dendritic cells in contact with primary CD4 T or B lymphocytes
- PMID: 20147388
- PMCID: PMC2863763
- DOI: 10.1128/JVI.01567-09
Stimulation of HIV-1 replication in immature dendritic cells in contact with primary CD4 T or B lymphocytes
Abstract
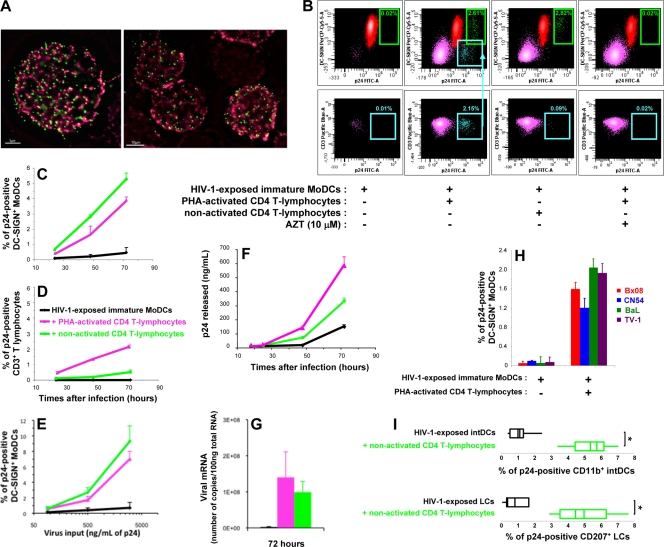
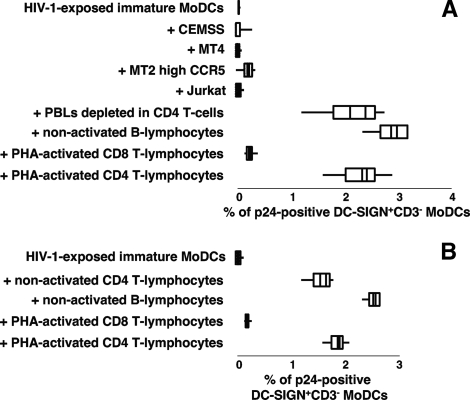
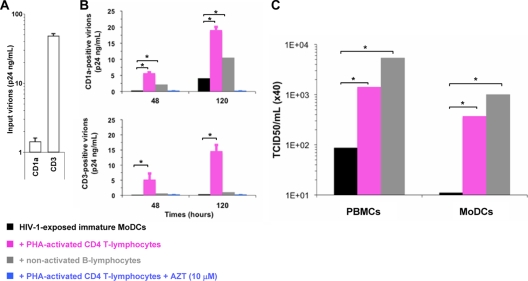
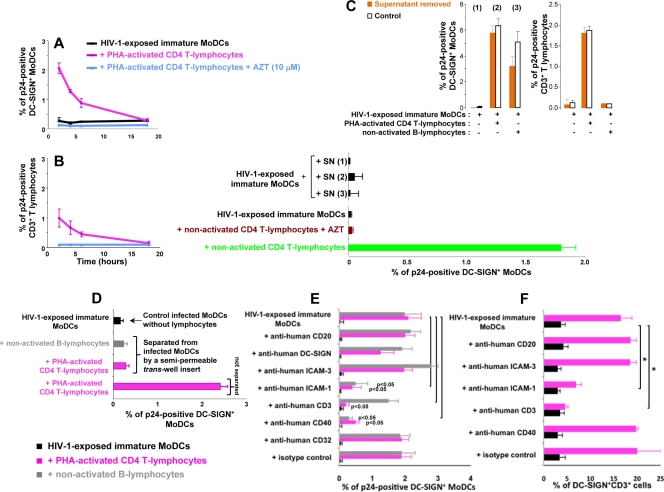
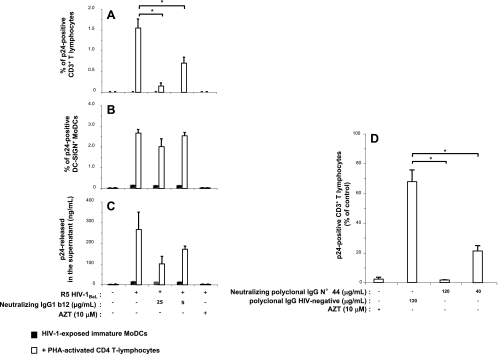
Sexual transmission is the major route of HIV-1 infection worldwide. Dendritic cells (DCs) from the mucosal layers are considered to be the initial targets of HIV-1 and probably play a crucial role in HIV-1 transmission. We investigated the role of cell-to-cell contact between HIV-1-exposed immature DCs and various lymphocyte subsets in the stimulation of HIV-1 replication. We found that HIV-1 replication and production in DCs were substantially enhanced by the coculture of DCs with primary CD4 T or nonpermissive B lymphocytes but not with primary activated CD8 T lymphocytes or human transformed CD4 T lymphocytes. Most of the new virions released by cocultures of HIV-1-exposed immature DCs and primary B lymphocytes expressed the DC-specific marker CD1a and were infectious for both immature DCs and peripheral blood mononuclear cells (PBMCs). Cocultured DCs thus produced large numbers of infectious viral particles under these experimental conditions. The soluble factors present in the supernatants of the cocultures were not sufficient to enhance HIV-1 replication in DCs, for which cell-to-cell contact was required. The neutralizing monoclonal antibody IgG1b12 and polyclonal anti-HIV-1 sera efficiently blocked HIV-1 transfer to CD4 T lymphocytes but did not prevent the increase in viral replication in DCs. Neutralizing antibodies thus proved to be more efficient at blocking HIV-1 transfer than previously thought. Our findings show that HIV-1 exploits DC-lymphocyte cross talk to upregulate replication within the DC reservoir. We provide evidence for a novel mechanism that may facilitate HIV-1 replication and transmission. This mechanism may favor HIV-1 pathogenesis, immune evasion, and persistence.
Figures
References
-
- Balkow, S., F. Krux, K. Loser, J. U. Becker, S. Grabbe, and U. Dittmer. 2007. Friend retrovirus infection of myeloid dendritic cells impairs maturation, prolongs contact to naive T cells, and favors expansion of regulatory T cells. Blood 110:3949-3958. - PubMed
-
- Banchereau, J., and R. M. Steinman. 1998. Dendritic cells and the control of immunity. Nature 392:245-252. - PubMed
-
- Brossard, C., V. Feuillet, A. Schmitt, C. Randriamampita, M. Romao, G. Raposo, and A. Trautmann. 2005. Multifocal structure of the T cell-dendritic cell synapse. Eur. J. Immunol. 35:1741-1753. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
