

Venezuelan equine Encephalitis virus capsid protein forms a tetrameric complex with CRM1 and importin alpha/beta that obstructs nuclear pore complex function
- PMID: 20147401
- PMCID: PMC2863722
- DOI: 10.1128/JVI.02554-09
Venezuelan equine Encephalitis virus capsid protein forms a tetrameric complex with CRM1 and importin alpha/beta that obstructs nuclear pore complex function
Abstract
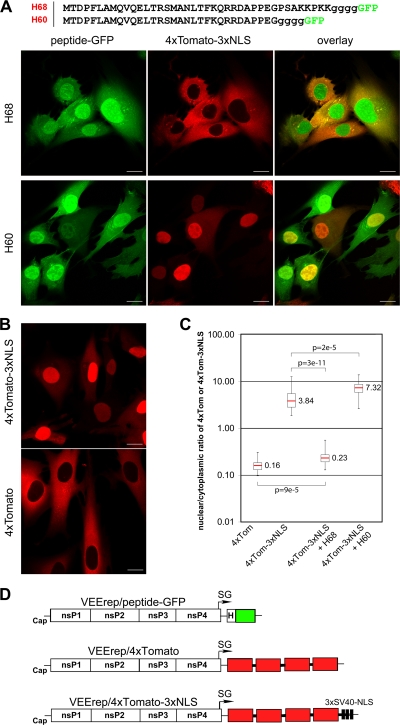
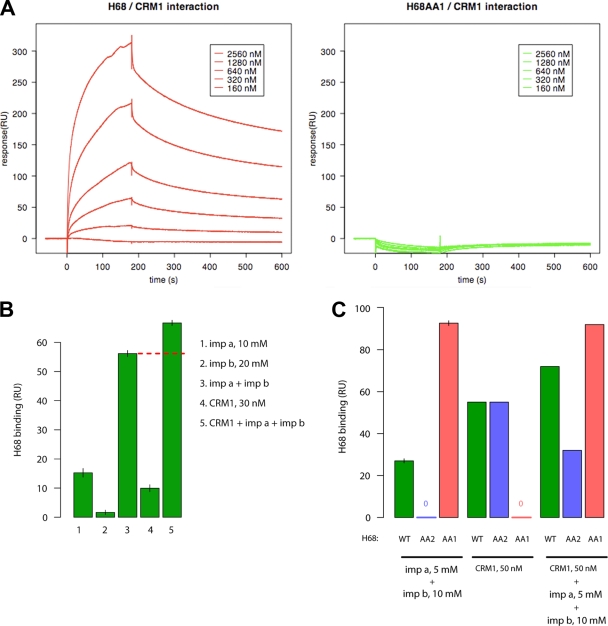
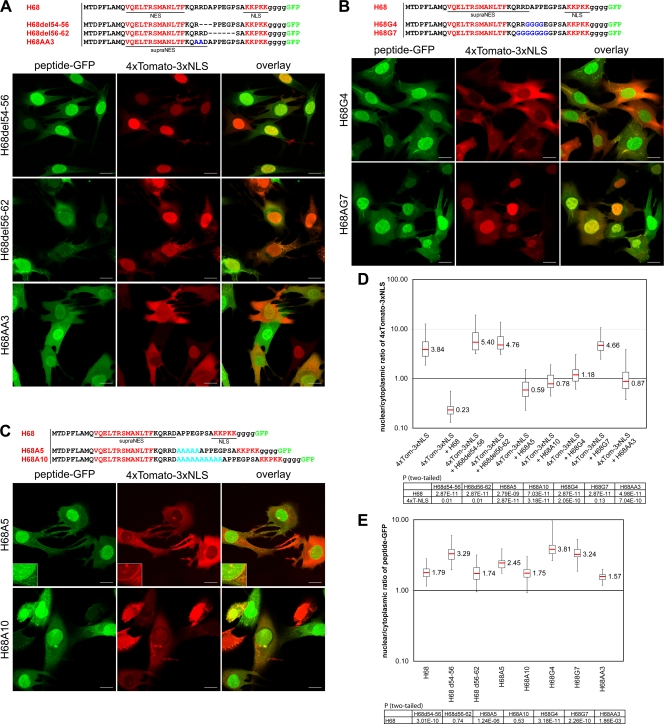
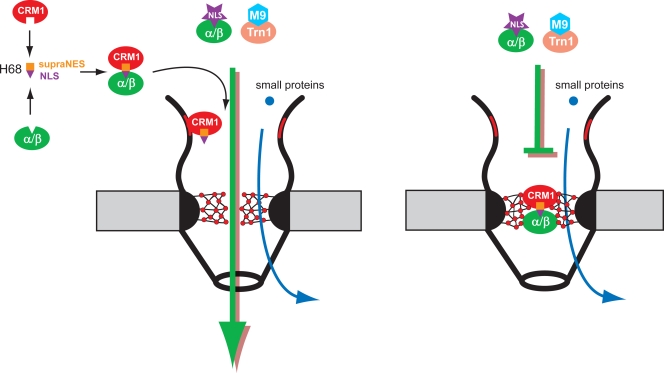
Development of the cellular antiviral response requires nuclear translocation of multiple transcription factors and activation of a wide variety of cellular genes. To counteract the antiviral response, several viruses have developed an efficient means of inhibiting nucleocytoplasmic traffic. In this study, we demonstrate that the pathogenic strain of Venezuelan equine encephalitis virus (VEEV) has developed a unique mechanism of nuclear import inhibition. Its capsid protein forms a tetrameric complex with the nuclear export receptor CRM1 and the nuclear import receptor importin alpha/beta. This unusual complex accumulates in the center channel of the nuclear pores and blocks nuclear import mediated by different karyopherins. The inhibitory function of VEEV capsid protein is determined by a short 39-amino-acid-long peptide that contains both nuclear import and supraphysiological nuclear export signals. Mutations in these signals or in the linker peptide attenuate or completely abolish capsid-specific inhibition of nuclear traffic. The less pathogenic VEEV strains contain a wide variety of mutations in this peptide that affect its inhibitory function in nuclear import. Thus, these mutations appear to be the determinants of this attenuated phenotype. This novel mechanism of inhibiting nuclear transport also shows that the nuclear pore complex is vulnerable to unusual cargo receptor complexes and sheds light on the importance of finely adjusted karyopherin-nucleoporin interactions for efficient cargo translocation.
Figures
References
-
- Aguilar, P. V., A. P. Adams, E. Wang, W. Kang, A. S. Carrara, M. Anishchenko, I. Frolov, and S. C. Weaver. 2008. Structural and nonstructural protein genome regions of eastern equine encephalitis virus are determinants of interferon sensitivity and murine virulence. J. Virol. 82:4920-4930. - PMC - PubMed
-
- Aguilar, P. V., I. P. Greene, L. L. Coffey, G. Medina, A. C. Moncayo, M. Anishchenko, G. V. Ludwig, M. J. Turell, M. L. O'Guinn, J. Lee, R. B. Tesh, D. M. Watts, K. L. Russell, C. Hice, S. Yanoviak, A. C. Morrison, T. A. Klein, D. J. Dohm, H. Guzman, A. P. Travassos da Rosa, C. Guevara, T. Kochel, J. Olson, C. Cabezas, and S. C. Weaver. 2004. Endemic Venezuelan equine encephalitis in northern Peru. Emerg. Infect. Dis. 10:880-888. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
