

Low pH-induced conformational change in herpes simplex virus glycoprotein B
- PMID: 20147407
- PMCID: PMC2849479
- DOI: 10.1128/JVI.02573-09
Low pH-induced conformational change in herpes simplex virus glycoprotein B
Abstract
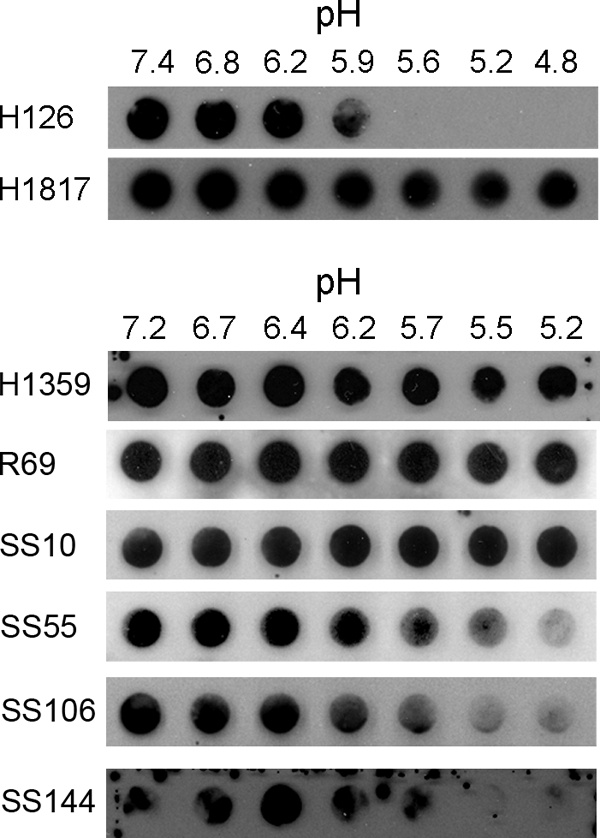
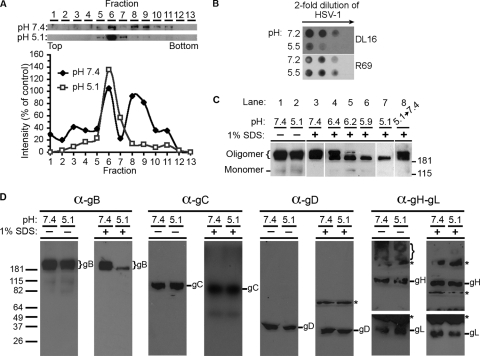
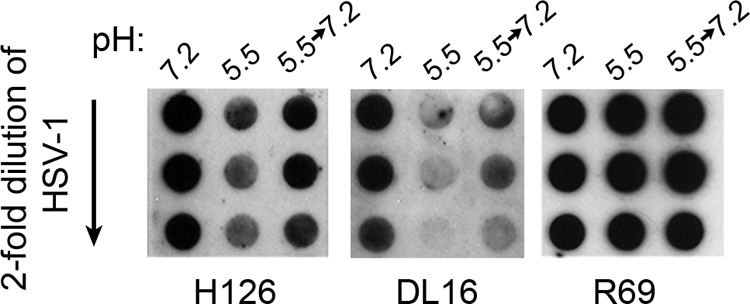
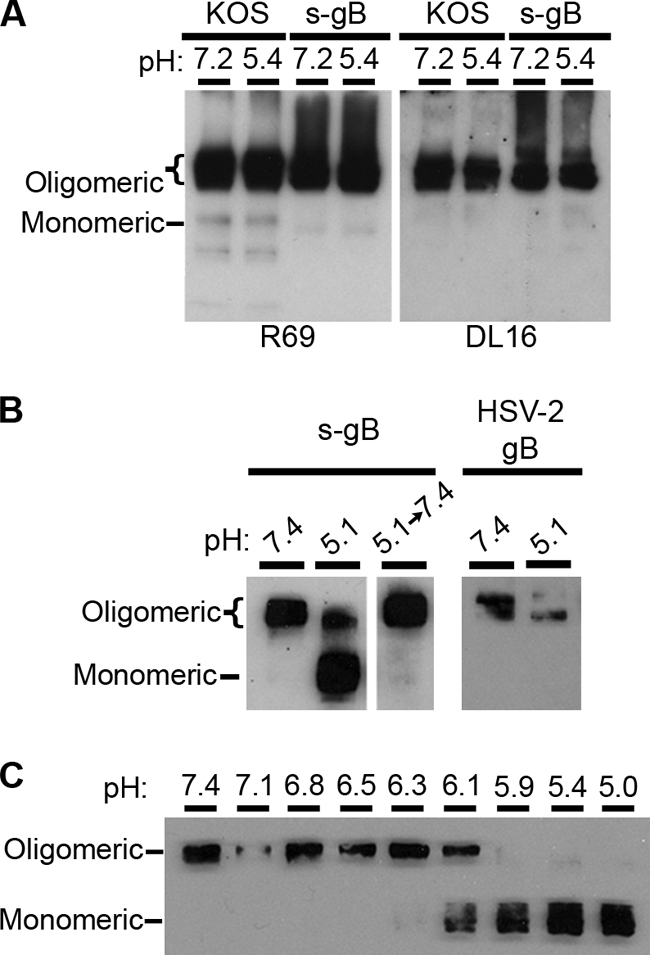
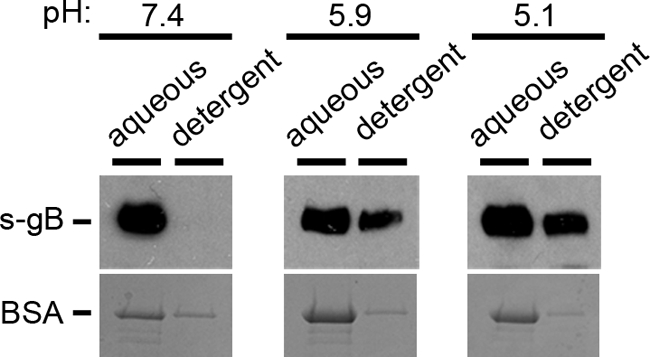
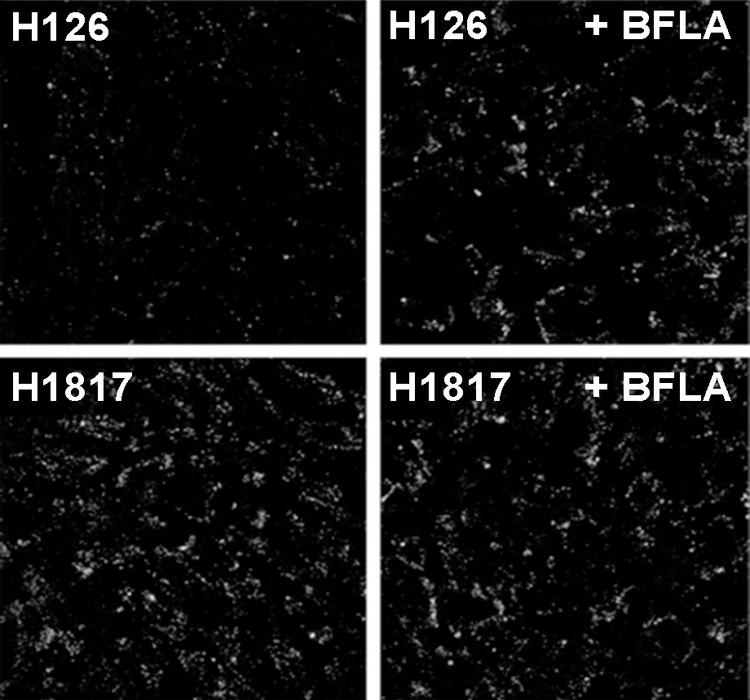
Herpesviruses can enter host cells using pH-dependent endocytosis pathways in a cell-specific manner. Envelope glycoprotein B (gB) is conserved among all herpesviruses and is a critical component of the complex that mediates membrane fusion and entry. Here we demonstrate that mildly acidic pH triggers specific conformational changes in herpes simplex virus (HSV) gB. The antigenic structure of gB was specifically altered by exposure to low pH both in vitro and during entry into host cells. The oligomeric conformation of gB was altered at a similar pH range. Exposure to acid pH appeared to convert virion gB into a lower-order oligomer. The detected conformational changes were reversible, similar to those in other class III fusion proteins. Exposure of purified, recombinant gB to mildly acidic pH resulted in similar changes in conformation and caused gB to become more hydrophobic, suggesting that low pH directly affects gB. We propose that intracellular low pH induces alterations in gB conformation that, together with additional triggers such as receptor binding, are essential for virion-cell fusion during herpesviral entry by endocytosis.
Figures
Similar articles
-
Mildly Acidic pH Triggers an Irreversible Conformational Change in the Fusion Domain of Herpes Simplex Virus 1 Glycoprotein B and Inactivation of Viral Entry.J Virol. 2017 Feb 14;91(5):e02123-16. doi: 10.1128/JVI.02123-16. Print 2017 Mar 1. J Virol. 2017. PMID: 28003487 Free PMC article.
-
Acidic pH Mediates Changes in Antigenic and Oligomeric Conformation of Herpes Simplex Virus gB and Is a Determinant of Cell-Specific Entry.J Virol. 2018 Aug 16;92(17):e01034-18. doi: 10.1128/JVI.01034-18. Print 2018 Sep 1. J Virol. 2018. PMID: 29925660 Free PMC article.
-
Reversible conformational change in herpes simplex virus glycoprotein B with fusion-from-without activity is triggered by mildly acidic pH.Virol J. 2010 Dec 1;7:352. doi: 10.1186/1743-422X-7-352. Virol J. 2010. PMID: 21122119 Free PMC article.
-
Low-pH-dependent changes in the conformation and oligomeric state of the prefusion form of herpes simplex virus glycoprotein B are separable from fusion activity.J Virol. 2011 Oct;85(19):9964-73. doi: 10.1128/JVI.05291-11. Epub 2011 Aug 3. J Virol. 2011. PMID: 21813610 Free PMC article.
-
Herpesvirus Entry into Host Cells Mediated by Endosomal Low pH.Traffic. 2016 Sep;17(9):965-75. doi: 10.1111/tra.12408. Epub 2016 May 24. Traffic. 2016. PMID: 27126894 Free PMC article. Review.
Cited by
-
Fusing structure and function: a structural view of the herpesvirus entry machinery.Nat Rev Microbiol. 2011 May;9(5):369-81. doi: 10.1038/nrmicro2548. Epub 2011 Apr 11. Nat Rev Microbiol. 2011. PMID: 21478902 Free PMC article. Review.
-
Capturing the herpes simplex virus core fusion complex (gB-gH/gL) in an acidic environment.J Virol. 2011 Jul;85(13):6175-84. doi: 10.1128/JVI.00119-11. Epub 2011 Apr 20. J Virol. 2011. PMID: 21507973 Free PMC article.
-
Impact of actin polymerization and filopodia formation on herpes simplex virus entry in epithelial, neuronal, and T lymphocyte cells.Front Cell Infect Microbiol. 2023 Nov 24;13:1301859. doi: 10.3389/fcimb.2023.1301859. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 38076455 Free PMC article. Review.
-
Towards Understanding KSHV Fusion and Entry.Viruses. 2019 Nov 18;11(11):1073. doi: 10.3390/v11111073. Viruses. 2019. PMID: 31752107 Free PMC article. Review.
-
Herpes simplex virus Membrane Fusion.Adv Anat Embryol Cell Biol. 2017;223:29-47. doi: 10.1007/978-3-319-53168-7_2. Adv Anat Embryol Cell Biol. 2017. PMID: 28528438 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
