

The effects of PKCalpha phosphorylation on the extensibility of titin's PEVK element
- PMID: 20149875
- PMCID: PMC2856808
- DOI: 10.1016/j.jsb.2010.02.002
The effects of PKCalpha phosphorylation on the extensibility of titin's PEVK element
Abstract
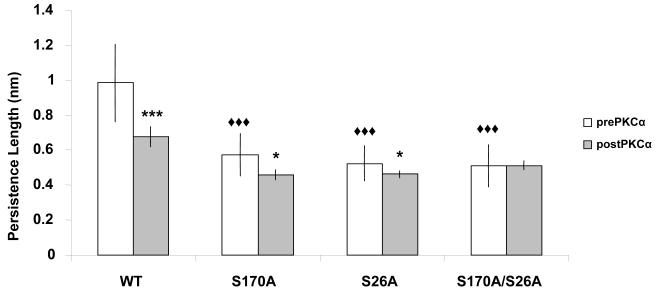
Post-translational modifications, along with isoform splicing, of titin determine the passive tension development of stretched sarcomeres. It was recently shown that PKCalpha phosphorylates two highly-conserved residues (S26 and S170) of the PEVK region in cardiac titin, resulting in passive tension increase. To determine how each phosphorylated residue affects myocardial stiffness, we generated three recombinant mutant PEVK fragments (S26A, S170A and S170A/S26A), each flanked by Ig domains. Single-molecule force spectroscopy shows that PKCalpha decreases the PEVK persistence length (from 0.99 to 0.68 nm); the majority of this decrease is attributable to phosphorylation of S26. Before PKCalpha, all three mutant PEVK fragments showed at least 40% decrease in persistence length compared to wildtype. Furthermore, Ig domain unfolding force measurements indicate that PEVK's flanking Ig domains are relatively unstable compared to other titin Ig domains. We conclude that phosphorylation of S26 is the primary mechanism through which PKCalpha modulates cardiac stiffness.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures





References
-
- Horowits R, et al. A physiological role for titin and nebulin in skeletal muscle. Nature. 1986;323(6084):160–4. - PubMed
-
- Labeit S, Kolmerer B. Titins: giant proteins in charge of muscle ultrastructure and elasticity. Science. 1995;270(5234):293–6. - PubMed
-
- Granzier HL, Labeit S. Titin and its associated proteins: the third myofilament system of the sarcomere. Adv Protein Chem. 2005;71:89–119. - PubMed
-
- Freiburg A, et al. Series of exon-skipping events in the elastic spring region of titin as the structural basis for myofibrillar elastic diversity. Circ Res. 2000;86(11):1114–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
