

AP-2alpha knockout mice exhibit optic cup patterning defects and failure of optic stalk morphogenesis
- PMID: 20150232
- PMCID: PMC2850623
- DOI: 10.1093/hmg/ddq060
AP-2alpha knockout mice exhibit optic cup patterning defects and failure of optic stalk morphogenesis
Abstract
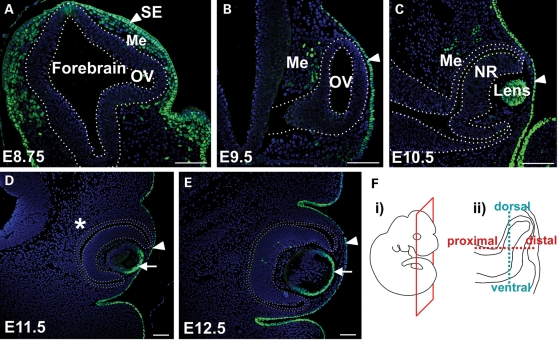
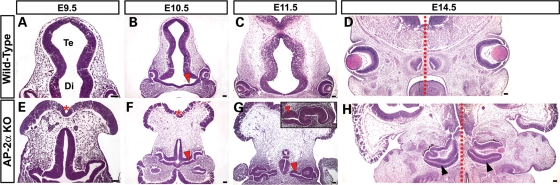
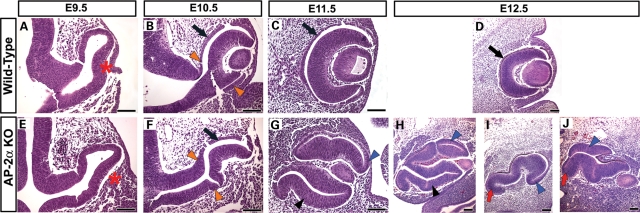
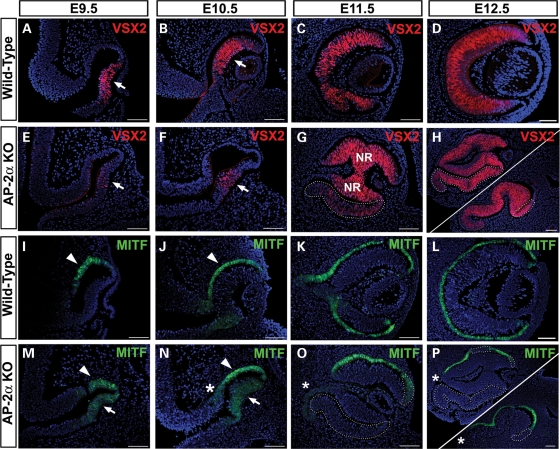
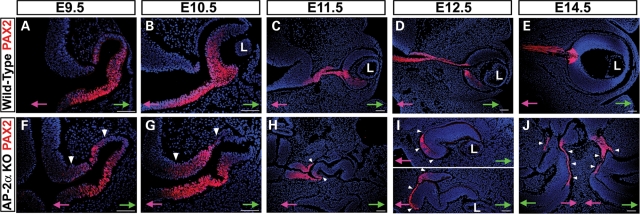
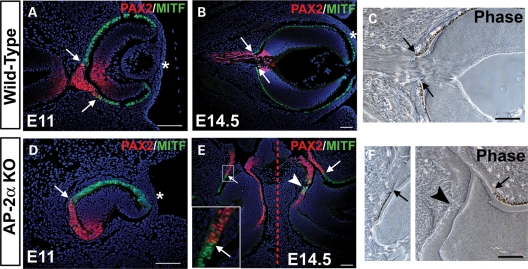
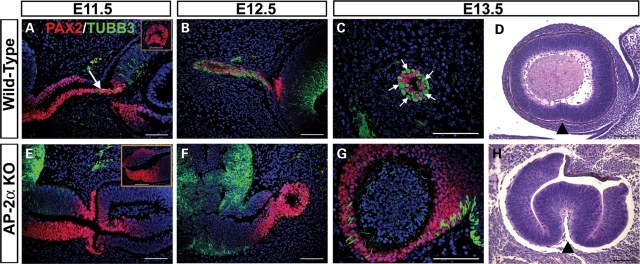
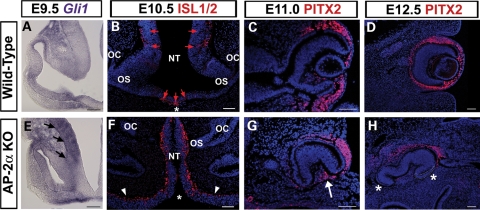
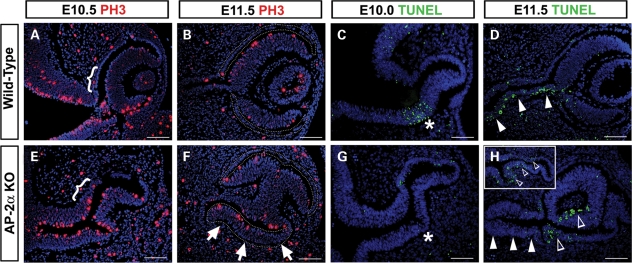
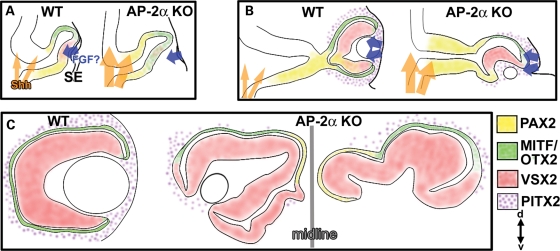
Appropriate development of the retina and optic nerve requires that the forebrain-derived optic neuroepithelium undergoes a precisely coordinated sequence of patterning and morphogenetic events, processes which are highly influenced by signals from adjacent tissues. Our previous work has suggested that transcription factor activating protein-2 alpha (AP-2alpha; Tcfap2a) has a non-cell autonomous role in optic cup (OC) development; however, it remained unclear how OC abnormalities in AP-2alpha knockout (KO) mice arise at the morphological and molecular level. In this study, we show that patterning and morphogenetic defects in the AP-2alpha KO optic neuroepithelium begin at the optic vesicle stage. During subsequent OC formation, ectopic neural retina and optic stalk-like tissue replaced regions of retinal pigment epithelium. AP-2alpha KO eyes also displayed coloboma in the ventral retina, and a rare phenotype in which the optic stalk completely failed to extend, causing the OCs to be drawn inward to the midline. We detected evidence of increased sonic hedgehog signaling in the AP-2alpha KO forebrain neuroepithelium, which likely contributed to multiple aspects of the ocular phenotype, including expansion of PAX2-positive optic stalk-like tissue into the OC. Our data suggest that loss of AP-2alpha in multiple tissues in the craniofacial region leads to severe OC and optic stalk abnormalities by disturbing the tissue-tissue interactions required for ocular development. In view of recent data showing that mutations in human TFAP2A result in similar eye defects, the current findings demonstrate that AP-2alpha KO mice provide a valuable model for human ocular disease.
Figures
References
-
- Rubenstein J.L., Beachy P.A. Patterning of the embryonic forebrain. Curr. Opin. Neurobiol. 1998;8:18–26. - PubMed
-
- Gage P.J., Rhoades W., Prucka S.K., Hjalt T. Fate maps of neural crest and mesoderm in the mammalian eye. Invest. Ophthalmol. Vis. Sci. 2005;46:4200–4208. - PubMed
-
- Chow R.L., Lang R.A. Early eye development in vertebrates. Annu. Rev. Cell. Dev. Biol. 2001;17:255–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
