

Transcriptional upregulation of both egl-1 BH3-only and ced-3 caspase is required for the death of the male-specific CEM neurons
- PMID: 20150917
- PMCID: PMC2902690
- DOI: 10.1038/cdd.2010.3
Transcriptional upregulation of both egl-1 BH3-only and ced-3 caspase is required for the death of the male-specific CEM neurons
Abstract
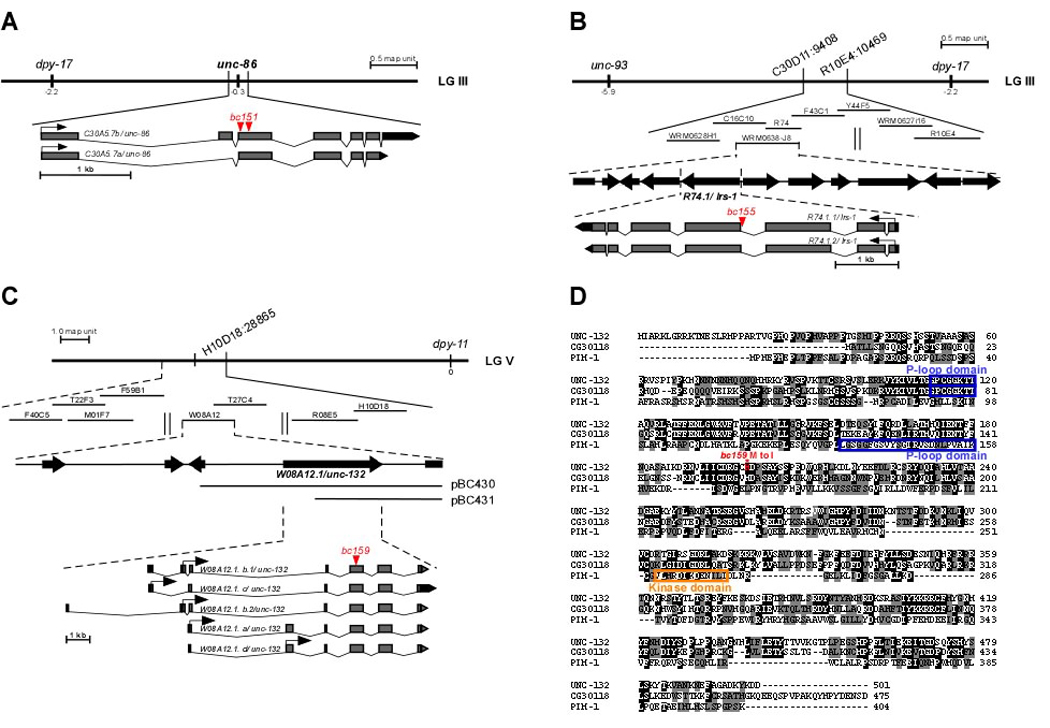
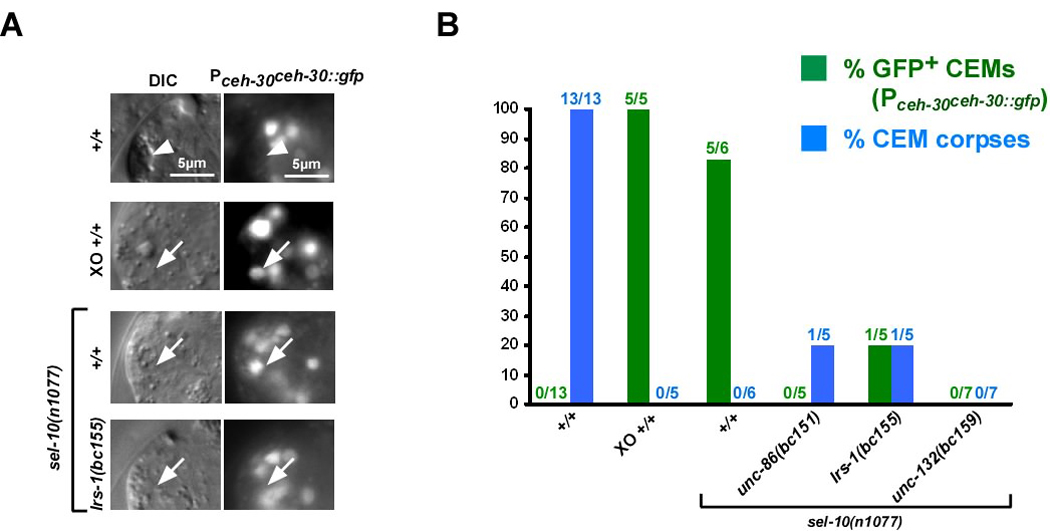
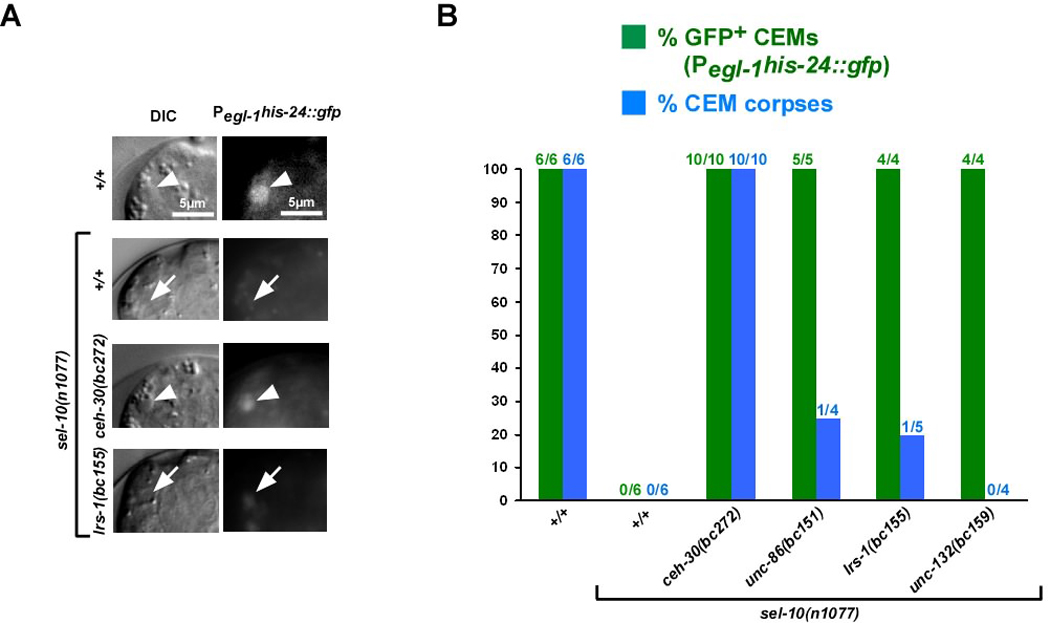
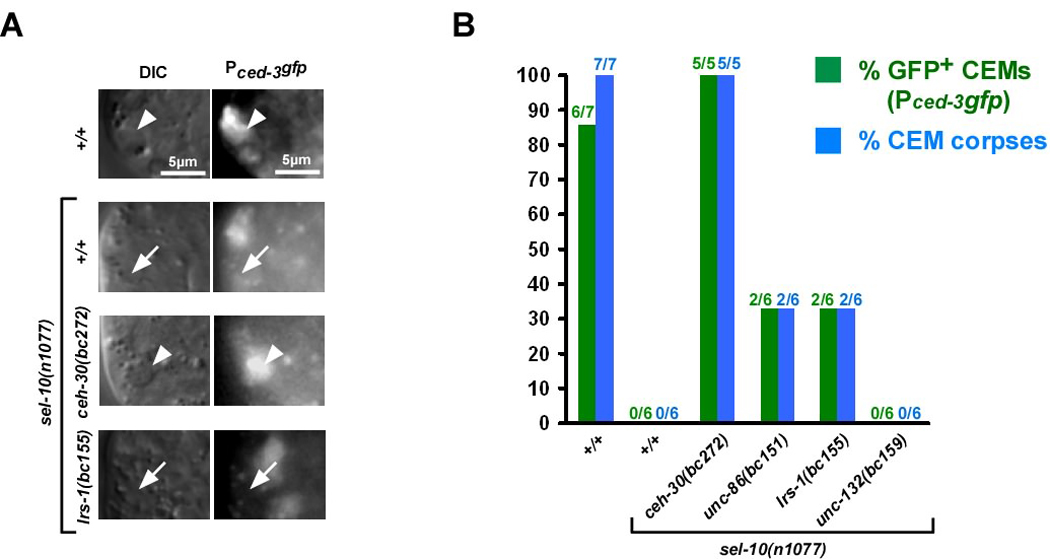
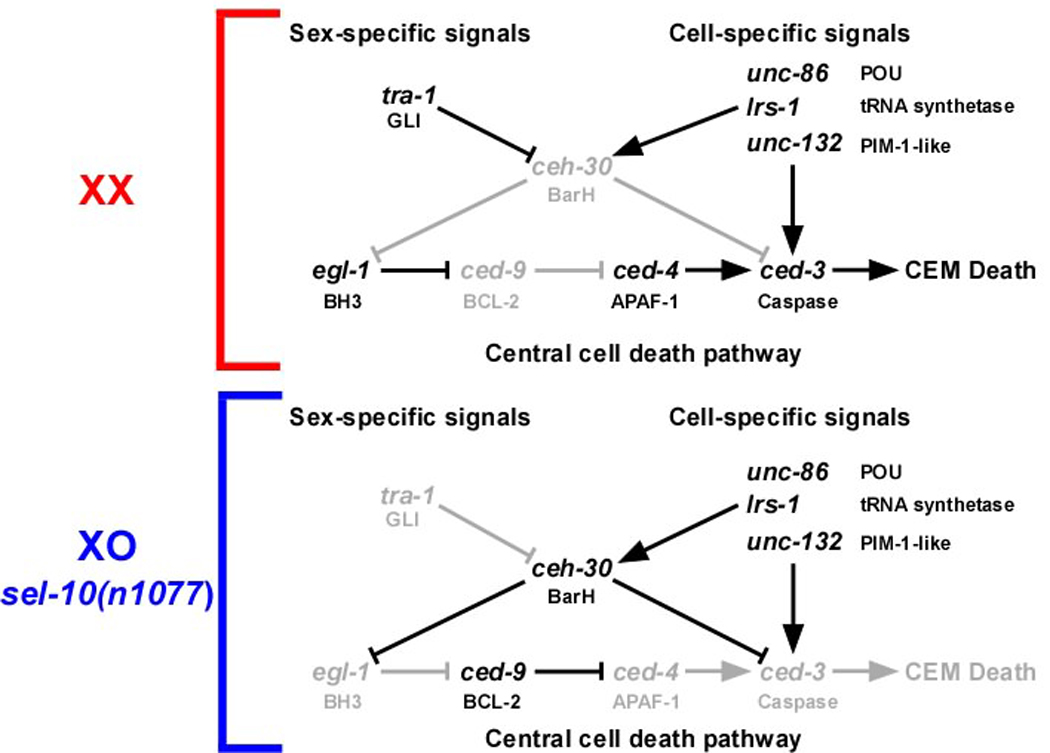
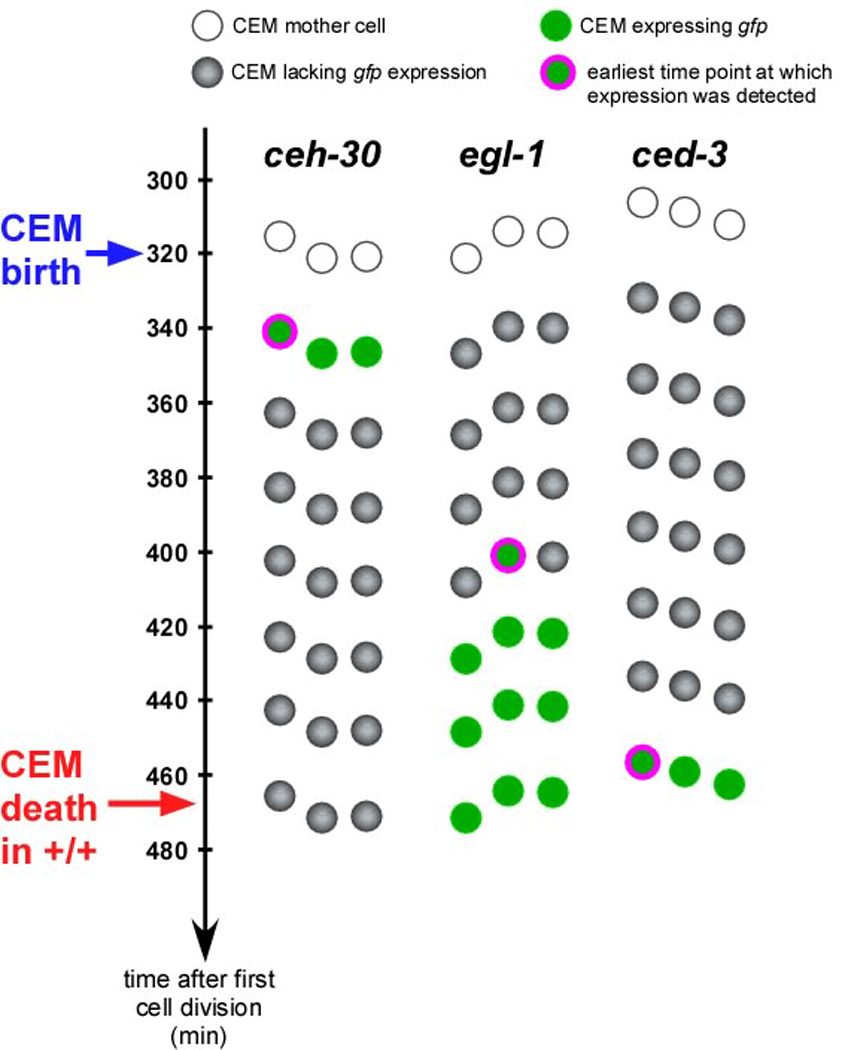
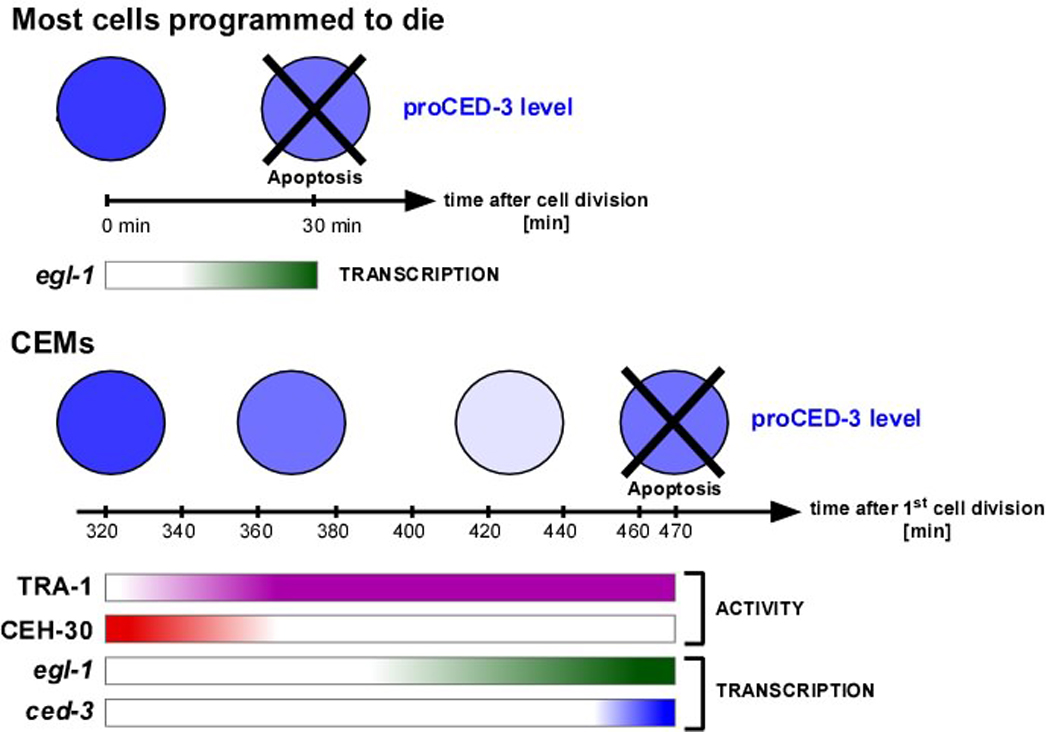
Most of the 131 cells that die during the development of a Caenorhabditis elegans hermaphrodite do so approximately 30 min after being generated. Furthermore, in these cells, the pro-caspase proCED-3 is inherited from progenitors and the transcriptional upregulation of the BH3-only gene egl-1 is thought to be sufficient for apoptosis induction. In contrast, the four CEM neurons, which die in hermaphrodites, but not males, die approximately 150 min after being generated. We found that in the CEMs, the transcriptional activation of both the egl-1 and ced-3 gene is necessary for apoptosis induction. In addition, we show that the Bar homeodomain transcription factor CEH-30 represses egl-1 and ced-3 transcription in the CEMs, thereby permitting their survival. Furthermore, we identified three genes, unc-86, lrs-1, and unc-132, which encode a POU homeodomain transcription factor, a leucyl-tRNA synthetase, and a novel protein with limited sequence similarity to the mammalian proto-oncoprotein and kinase PIM-1, respectively, that promote the expression of the ceh-30 gene in the CEMs. On the basis of these results, we propose that egl-1 and ced-3 transcription are coregulated in the CEMs to compensate for limiting proCED-3 levels, which most probably are a result of proCED-3 turn over. Similar coregulatory mechanisms for BH3-only proteins and pro-caspases may function in higher organisms to allow efficient apoptosis induction. Finally, we present evidence that the timing of the death of the CEMs is controlled by TRA-1 Gli, the terminal global regulator of somatic sexual fate in C. elegans.
Figures
Similar articles
-
Control of sex-specific apoptosis in C. elegans by the BarH homeodomain protein CEH-30 and the transcriptional repressor UNC-37/Groucho.Genes Dev. 2007 Dec 1;21(23):3195-207. doi: 10.1101/gad.1607807. Genes Dev. 2007. PMID: 18056429 Free PMC article.
-
The C. elegans protein CEH-30 protects male-specific neurons from apoptosis independently of the Bcl-2 homolog CED-9.Genes Dev. 2007 Dec 1;21(23):3181-94. doi: 10.1101/gad.1607007. Genes Dev. 2007. PMID: 18056428 Free PMC article.
-
Timing of the onset of a developmental cell death is controlled by transcriptional induction of the C. elegans ced-3 caspase-encoding gene.Development. 2007 Apr;134(7):1357-68. doi: 10.1242/dev.02818. Epub 2007 Feb 28. Development. 2007. PMID: 17329362
-
Noncanonical cell death programs in the nematode Caenorhabditis elegans.Cell Death Differ. 2008 Jul;15(7):1124-31. doi: 10.1038/cdd.2008.56. Epub 2008 Apr 25. Cell Death Differ. 2008. PMID: 18437162 Review.
-
Cell death specification in C. elegans.Cell Cycle. 2008 Aug 15;7(16):2479-84. doi: 10.4161/cc.7.16.6479. Epub 2008 Aug 19. Cell Cycle. 2008. PMID: 18719375 Free PMC article. Review.
Cited by
-
Repurposing the Killing Machine: Non-canonical Roles of the Cell Death Apparatus in Caenorhabditis elegans Neurons.Front Cell Dev Biol. 2022 Feb 14;10:825124. doi: 10.3389/fcell.2022.825124. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35237604 Free PMC article. Review.
-
T-Cell Factors as Transcriptional Inhibitors: Activities and Regulations in Vertebrate Head Development.Front Cell Dev Biol. 2021 Nov 24;9:784998. doi: 10.3389/fcell.2021.784998. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34901027 Free PMC article. Review.
-
Cell Death in C. elegans Development.Curr Top Dev Biol. 2015;114:1-42. doi: 10.1016/bs.ctdb.2015.07.018. Epub 2015 Sep 9. Curr Top Dev Biol. 2015. PMID: 26431562 Free PMC article. Review.
-
Activity-Dependent Regulation of the Proapoptotic BH3-Only Gene egl-1 in a Living Neuron Pair in Caenorhabditis elegans.G3 (Bethesda). 2019 Nov 5;9(11):3703-3714. doi: 10.1534/g3.119.400654. G3 (Bethesda). 2019. PMID: 31519744 Free PMC article.
-
A genetic screen identifies C. elegans eif-3.H and hrpr-1 as pro-apoptotic genes and potential activators of egl-1 expression.MicroPubl Biol. 2024 Feb 16;2024:10.17912/micropub.biology.001126. doi: 10.17912/micropub.biology.001126. eCollection 2024. MicroPubl Biol. 2024. PMID: 38434221 Free PMC article.
References
-
- Vaux DL, Korsmeyer SJ. Cell death in development. Cell. 1999;96(2):245–254. - PubMed
-
- Adams JM. Ways of dying: multiple pathways to apoptosis. Genes Dev. 2003 Oct 15;17(20):2481–2495. - PubMed
-
- Danial NN, Korsmeyer SJ. Cell death: critical control points. Cell. 2004 Jan 23;116(2):205–219. - PubMed
-
- Horvitz HR. Nobel lecture. Worms, life and death. Biosci Rep. 2003 Oct-Dec;23(5–6):239–303. - PubMed
-
- Lettre G, Hengartner MO. Developmental apoptosis in C. elegans: a complex CEDnario. Nature Reviews Molecular Cell Biology. 2006;7:97–108. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
