

Reduction of oxidative cellular damage by overexpression of the thioredoxin TRX2 gene improves yield and quality of wine yeast dry active biomass
- PMID: 20152017
- PMCID: PMC2835662
- DOI: 10.1186/1475-2859-9-9
Reduction of oxidative cellular damage by overexpression of the thioredoxin TRX2 gene improves yield and quality of wine yeast dry active biomass
Erratum in
- Microb Cell Fact. 2012;11:31
Abstract
Background: Wine Saccharomyces cerevisiae strains, adapted to anaerobic must fermentations, suffer oxidative stress when they are grown under aerobic conditions for biomass propagation in the industrial process of active dry yeast production. Oxidative metabolism of sugars favors high biomass yields but also causes increased oxidation damage of cell components. The overexpression of the TRX2 gene, coding for a thioredoxin, enhances oxidative stress resistance in a wine yeast strain model. The thioredoxin and also the glutathione/glutaredoxin system constitute the most important defense against oxidation. Trx2p is also involved in the regulation of Yap1p-driven transcriptional response against some reactive oxygen species.
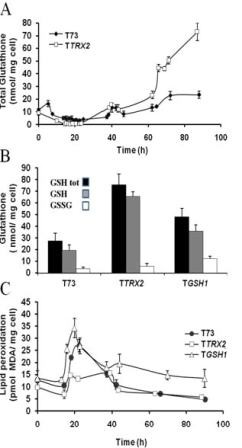
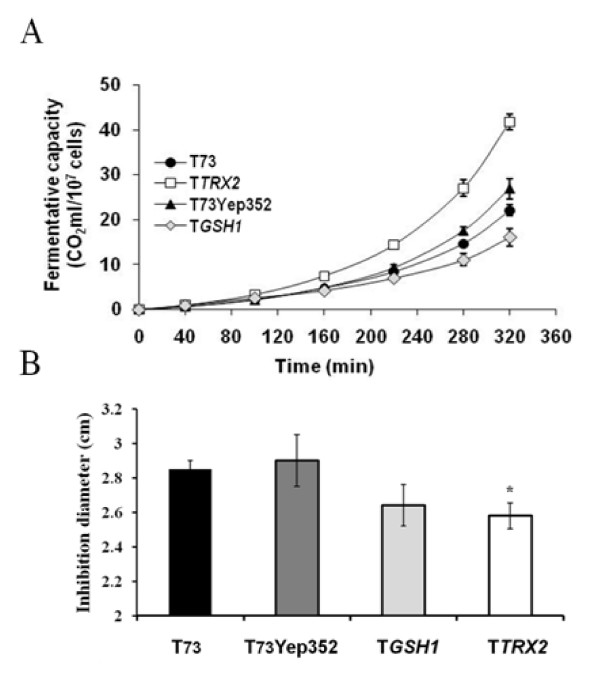
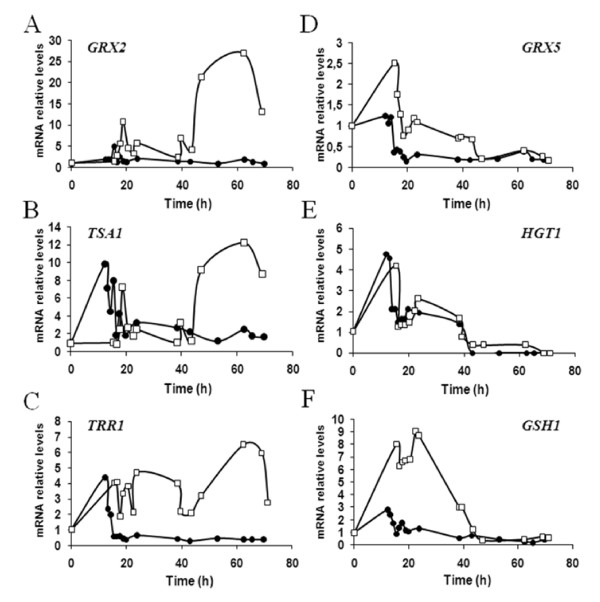
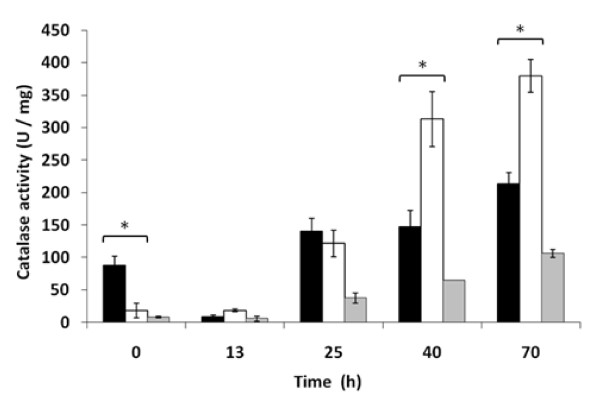
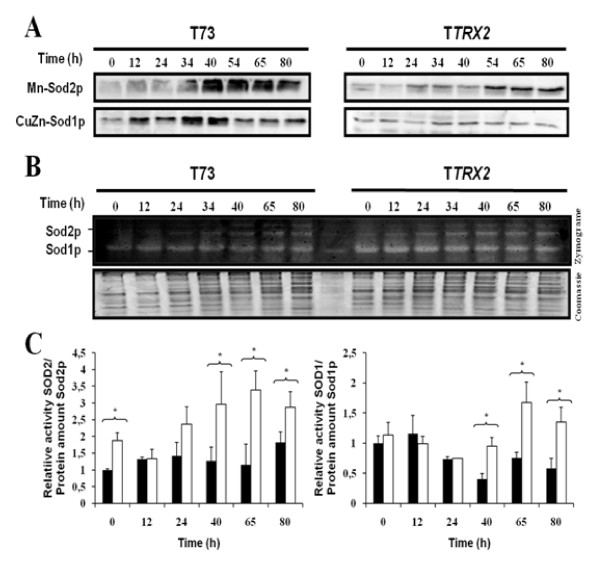
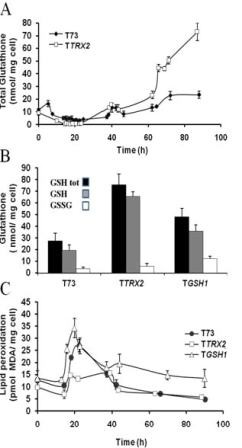
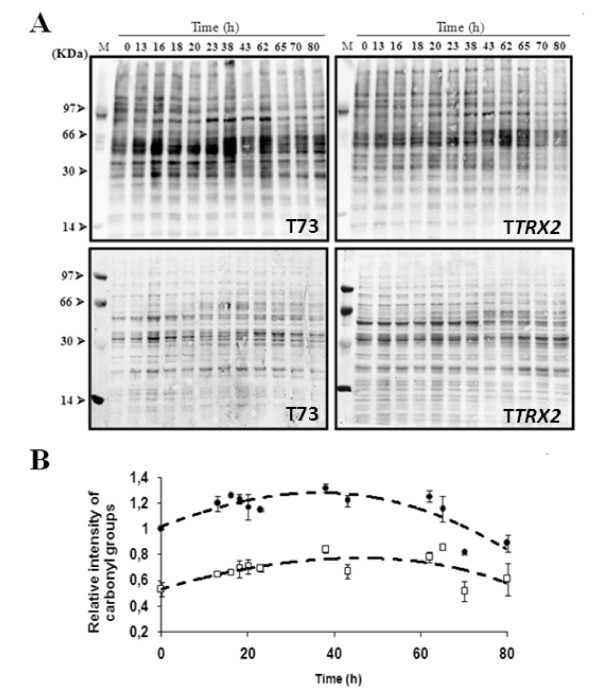
Results: Laboratory scale simulations of the industrial active dry biomass production process demonstrate that TRX2 overexpression increases the wine yeast final biomass yield and also its fermentative capacity both after the batch and fed-batch phases. Microvinifications carried out with the modified strain show a fast start phenotype derived from its enhanced fermentative capacity and also increased content of beneficial aroma compounds. The modified strain displays an increased transcriptional response of Yap1p regulated genes and other oxidative stress related genes. Activities of antioxidant enzymes like Sod1p, Sod2p and catalase are also enhanced. Consequently, diminished oxidation of lipids and proteins is observed in the modified strain, which can explain the improved performance of the thioredoxin overexpressing strain.
Conclusions: We report several beneficial effects of overexpressing the thioredoxin gene TRX2 in a wine yeast strain. We show that this strain presents an enhanced redox defense. Increased yield of biomass production process in TRX2 overexpressing strain can be of special interest for several industrial applications.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
