

Review
doi: 10.1016/j.cub.2009.10.023.
Common mechanisms of synaptic plasticity in vertebrates and invertebrates
Affiliations
- PMID: 20152143
- PMCID: PMC5125775
- DOI: 10.1016/j.cub.2009.10.023
Item in Clipboard
Review
Common mechanisms of synaptic plasticity in vertebrates and invertebrates
Curr Biol.
.
Abstract
Until recently, the literature on learning-related synaptic plasticity in invertebrates has been dominated by models assuming plasticity is mediated by presynaptic changes, whereas the vertebrate literature has been dominated by models assuming it is mediated by postsynaptic changes. Here I will argue that this situation does not reflect a biological reality and that, in fact, invertebrate and vertebrate nervous systems share a common set of mechanisms of synaptic plasticity.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures
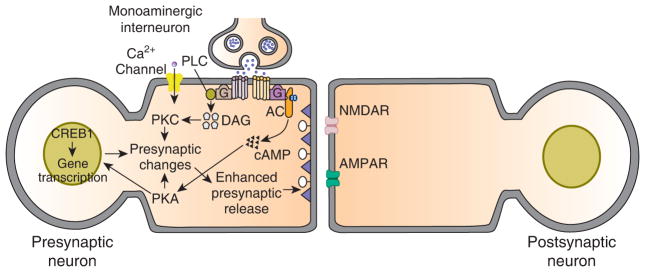
General presynaptic model for learning-related synaptic enhancement in invertebrates circa 1995. This model is a hybrid and is based on results from cellular and molecular work on Aplysia and the leech, and genetic work on Drosophila. The main basis for plasticity is heterosynaptic modulatory input from monoaminergic interneurons. In Aplysia, as well as in the leech [48], at least some of the modulatory interneurons contain 5-HT, whereas in Drosophila the transmitter used by the modulatory interneurons is dopamine or octopamine [49]. According to the model, the monoamine binds to G protein-coupled receptors on the presynaptic neuron, and stimulates the activity of several kinases: prominent among these is PKA; presynaptic PKC has also been implicated in synaptic facilitation in Aplysia, and has been shown to play a role in associative learning in Drosophila. In Aplysia, PKA and PKC phosphorylate substrate proteins that lead to closure of K+ channels, as well as enhance mobilization of presynaptic vesicles. Pprolonged activation of PKA results in its translocation to the nucleus of the presynaptic cell, where it activates the transcription factor CREB1, and thereby triggers long-term (≥ 24 hour) changes. Activation of CREB1 involves its release from repression by the inhibitory isoform CREB2 in Aplysia and Drosophila; once activated, CREB1 stimulates gene transcription. Some of the induce genes themselves encode transcription factors, such as C/EBP. The products of the gene transcription mediate a variety of long-term cellular changes, including persistent closure of ion channels and growth of new presynaptic varicosities. The basic model illustrated represents a cellular mechanism for sensitization, a nonassociative form of memory. The model originally proposed to explain classical conditioning in invertebrates is a modification of the presynaptic model for sensitization. According to the original presynaptic model, delivery of the conditioned stimulus (siphon touch in Aplysia) causes an influx of Ca2+ into the neuron through voltage-dependent Ca2+ channels. The combined Ca2+/calmodulin activates the adenylyl cyclase, which can be stimulated by both Ca2+/calmodulin and G-protein activation via the monoamine. Release of the monoamine is produced by the unconditioned stimulus (tail shock in Aplysia). Paired delivery of the conditioned and unconditioned stimuli results in dual stimulation of the cyclase by Ca2+/calmodulin and the monoamine. This dual stimulation of the cyclase yields greater synthesis of cAMP and, hence, greater activation of PKA, than either Ca2+/calmodulin or the monoamine alone. This dual stimulation of the presynaptic adenylyl cyclase, according to the model, is the basis for associative synaptic enhancement. In the case of Aplysia the presynaptic neuron is a central mechanosensory neuron, whereas the postsynaptic neuron is a central motor neurons. In the leech the presynaptic neuron is also a central mechanosensory neuron, and the postsynaptic neuron is the S-cell interneuron [24]. In Drosophila the presynaptic neuron is thought to be a mushroom body Kenyon cell [49].
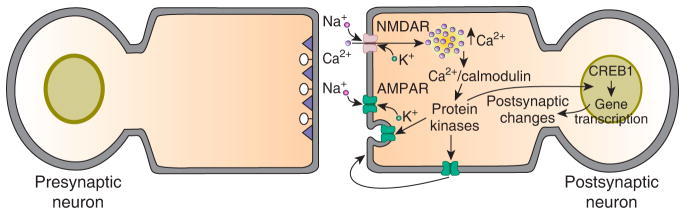
Standard model for NMDA receptor-dependent LTP in the mammalian hippocampus and cortex. There are two phases to this form of LTP, an early phase that lasts for one to two hours and a late phase that can last for eight hours or more in slice preparations. In the model activation of postsynaptic NMDA receptors, either by high-frequency stimulation or paired pre- and postsynaptic (or Hebbian) stimulation, produces an influx of Ca2+ into postsynaptic dendrites via the open NMDA receptor channels. This postsynaptic influx of Ca2+ activates several kinases that are involved in the induction and expression of LTP. Among these kinases are Ca2+/calmodulin-dependent protein kinase II (CaMKII), PKA, mitogen-activated protein kinases (MAPK) and protein kinase M zeta (PKMζ) [20]. Activation of these kinases causes insertion of new AMPA receptors into the postsynaptic membrane, which is believed to be the major expression mechanism for this form of synaptic plasticity. The mode of AMPA receptor insertion is controversial; the main competing schemes are lateral diffusion of extrasynaptic receptors into postsynaptic sites [22] and exocytotic insertion of the receptors into the postsynaptic membrane [20]. Prolonged activation of the postsynaptic kinases, such as occurs with the multiple, spaced trains of high-frequency stimulation that are used to induce late-LTP, cause gene transcription and protein synthesis, both of which are required for late-LTP. There is evidence for a role for CREB1-dependent transcription in late-LTP. The gene products are believed to be involved in structural remodeling of the postsynaptic neuron, particularly the growth and stabilization of new dendritic spines.
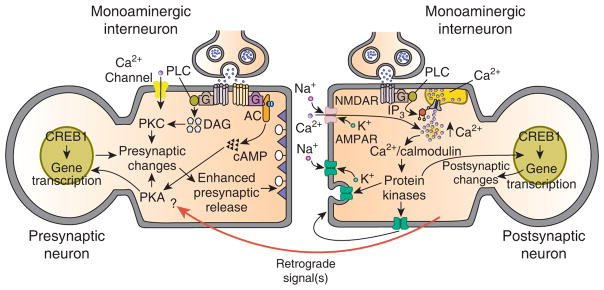
General model for learning-related enhancement of excitatory glutamatergic synapses. This model is based on recent data from studies of synaptic plasticity in invertebrates and vertebrates. Two prominent features of the model are postsynaptic modulatory input from monoaminergic interneurons and retrograde signaling. Evidence for a postsynaptic contribution of monoamines comes from recent studies in Aplysia [32,33] and mammals [50]. In Aplysia, prolonged stimulation with 5-HT causes modulation of AMPA receptor trafficking, which appears to involve exocytotic insertion of AMPA receptors into the cell membrane of the motor neuron. This process is mediated by G protein-stimulated release of Ca2+ from intracellular stores. In the mammalian hippocampus there is evidence that heterosynaptic modulatory input from the basolateral amygdala can convert early-LTP to late-LTP [50]. In this instance, the heterosynaptic input may be from norepinephrine- or acetylcholine-containing axons. In other instances, the heterosynaptic modulatory input may be from peptidergic axons as well. (Only monoaminergic interneurons are shown in the model for the purpose of simplicity.) As shown in the model, the conversion of early-LTP to late-LTP could be mediated by the summing of separate pools of intracellular Ca2+, one resulting from open NMDA receptor channels and the other from release of Ca2+ from intracellular stores, which is stimulated by the heterosynaptic monoaminergic input. The elevated intracellular Ca2+ is also responsible, either directly or indirectly (perhaps through protein synthesis), for triggering the activation of one or more retrograde signals; the retrograde signals, in turn, contribute critically to presynaptic changes and enhanced presynaptic release [41]. An important question is the identity of the presynaptic molecules that are the targets of the retrograde signals. In Aplysia there is evidence that one of these presynaptic targets may be PKA [37,42]. Another important question is to what extent the long-term presynaptic changes result from an interaction between the heterosynaptic input to the presynaptic neuron and the retrograde signal(s). An intriguing possibility is that retrograde signaling alone may be sufficient for long-term changes in the presynaptic neuron.
Similar articles
-
Regulation of synaptic function by neurotrophic factors in vertebrates and invertebrates: implications for development and learning.Learn Mem. 1999 May-Jun;6(3):193-215. Learn Mem. 1999. PMID: 10492003 Review.
-
On the quantal analysis of hippocampal long-term potentiation and related phenomena of synaptic plasticity.Neuroscience. 1993 Sep;56(2):275-304. doi: 10.1016/0306-4522(93)90332-a. Neuroscience. 1993. PMID: 8247261 Review. No abstract available.
-
Tyrosine kinases, synaptic plasticity and memory: insights from vertebrates and invertebrates.Trends Neurosci. 2003 Nov;26(11):625-30. doi: 10.1016/j.tins.2003.09.005. Trends Neurosci. 2003. PMID: 14585603 Review.
-
Synaptic Transmission Optimization Predicts Expression Loci of Long-Term Plasticity.Neuron. 2017 Sep 27;96(1):177-189.e7. doi: 10.1016/j.neuron.2017.09.021. Neuron. 2017. PMID: 28957667 Free PMC article.
-
Learning-related synaptic plasticity: LTP and LTD.Curr Opin Neurobiol. 1991 Jun;1(1):113-20. doi: 10.1016/0959-4388(91)90018-3. Curr Opin Neurobiol. 1991. PMID: 1822291 Review.
Cited by
-
Activity-dependent plasticity of spinal circuits in the developing and mature spinal cord.Neural Plast. 2012;2012:964843. doi: 10.1155/2012/964843. Epub 2012 Aug 1. Neural Plast. 2012. PMID: 22900208 Free PMC article. Review.
-
Neurobiological reduction: From cellular explanations of behavior to interventions.Front Psychol. 2022 Dec 22;13:987101. doi: 10.3389/fpsyg.2022.987101. eCollection 2022. Front Psychol. 2022. PMID: 36619115 Free PMC article. Review.
-
Proteomic Analysis of Brain Region and Sex-Specific Synaptic Protein Expression in the Adult Mouse Brain.Cells. 2020 Jan 28;9(2):313. doi: 10.3390/cells9020313. Cells. 2020. PMID: 32012899 Free PMC article.
-
The biopsychology of salt hunger and sodium deficiency.Pflugers Arch. 2015 Mar;467(3):445-56. doi: 10.1007/s00424-014-1676-y. Epub 2015 Jan 10. Pflugers Arch. 2015. PMID: 25572931 Free PMC article. Review.
-
Mapping molecular memory: navigating the cellular pathways of learning.Cell Mol Neurobiol. 2012 Aug;32(6):919-41. doi: 10.1007/s10571-012-9836-0. Epub 2012 Apr 10. Cell Mol Neurobiol. 2012. PMID: 22488526 Free PMC article. Review.
References
-
- Castellucci V, Pinsker H, Kupfermann I, Kandel ER. Neuronal mechanisms of habituation and dishabituation of the gill-withdrawal reflex in Aplysia. Science. 1970;167:1745–1748. - PubMed
-
- Castellucci VF, Kandel ER. Presynaptic facilitation as a mechanism for behavioral sensitization in Aplysia. Science. 1976;194:1176–1178. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
